
ESBE ARA659 45/120сек. 6Нм пропорциональный электропривод для клапанов смесит.
Увеличить
Акция «Вместе в соцсетях»: вы сэкономите: 201 грн
Расскажите друзьям в социальной сети про этот товар, сообщите нам адрес своего !
Цена : 6,701 грн
Нашли дешевле?
Моментальный заказ
Укажите номер телефона, и менеджер позвонит Вам
Ваш заказ успешно оформлен, менеджер в ближайшее время перезвонит Вам, для уточнения деталей заказа.
Что-то пошло не так, обновите страницу.
Найти монтажника в своем городе Обсудить товар на форуме
Способы доставки
- Доставка по Украине в любой населенный пункт службами “Новая Почта”, “Укрпочта”, “Міст Експрес” и др.
- Доставка по Львову – курьером
Способы оплаты
Подробнее
- Наличная при доставке курьером
- Наложенным платежом при получении заказа
- Описание полное
- Описание товара
- Характеристики
- Документация
- Отзывы (1) Array
- Видео покупателей (0)
Описание ESBE ARA659 45/120сек.
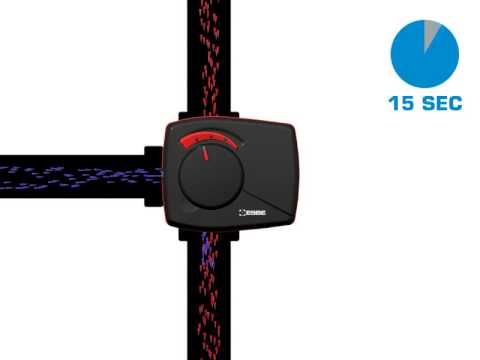 6Нм пропорциональный электропривод для клапанов смесит.
6Нм пропорциональный электропривод для клапанов смесит.РОТАЦИОННЫЕ МОТОРИЗОВАННЫЕ КЛАПАНЫ
ПРИВОД
СЕРИЯ ARA600
Приводы ESBE серии ARA600 для управления смесительными клапанами ESBE DN 15–50. Приводы имеют диапазон действия 90° и могут легко управляться вручную.Компактный пропорциональный привод ARA659 (24В переменного/постоянного тока) сконструирован для управления ротационным смесительным клапаном DN 15-50. Привод управляется пропорциональным сигналом и рекомендуется для установки на смесительный клапан. Привод имеет диапазон действия 90° и клапан легко может управляться вручную при помощи рукоятки “нажать и повернуть” на передней части привода
Особенностью данного типа приводов является возможность передачи выходного сигнала для устройств мониторинга и управления. Также электропривод оснащён устройством, снижающим помехи входного сигнала и устройством запоминания рабочей позиции на случай временного отключения питания.
ПОДХОДЯЩИЕ СМЕСИТЕЛЬНЫЕ КЛАПАНЫ
Благодаря специальному соединению между приводом серий ARA600 и клапанами ESBE серий VRG100, VRG200 и VRB100, устройство при регулировании в целом имеет уникальную точность и устойчивость. Привод серии ARA600 также легко устанавливать на клапанах ESBE серий MG, G, F, BIV, H и HG.
- Серия VRG100
- Серия VRG200
- Серия VRG300
- Серия VRB100
- Серия MG
- Серия G
- Серия F ≤ DN50
- Серия BIV
- Серия H и HG
ПРИСОЕДИНИТЕЛЬНЫЕ КОМПЛЕКТЫ
Привод поставляется вместе с переходником для облегчения подключения к ротационному смесительному клапану ESBE. Переходники могут быть заказаны отдельно.
Характеристики ESBE ARA659 45/120сек. 6Нм пропорциональный электропривод для клапанов смесит.
Номинальное потребление тока (A) | 8/4 |
|---|---|
Электрическое питание (В/Гц) | 24 ±10% В, 50/60 Гц |
Управляющий сигнал | 0. .10 В, 2..10 В, 0..20 мА, 4..20 мА .10 В, 2..10 В, 0..20 мА, 4..20 мА |
Время закрытия 90° (s) | 45/120 |
Крутящий момент (Нм) | 6 |
Величина вспомогательного выключателя | 6(3)А 250 В переменного тока |
Вес (кг) | 0,4 |
Степень защиты | IP41 |
Температура окружающей среды, °C | от -5 до +55 |
Отзывы о товаре
РОТАЦИОННЫЕ МОТОРИЗОВАННЫЕ КЛАПАНЫ
ПРИВОД
СЕРИЯ ARA600
Приводы ESBE серии ARA600 для управления смесительными клапанами ESBE DN 15–50. Приводы имеют диапазон действия 90° и могут легко управляться вручную.
Компактный пропорциональный привод ARA659 (24В переменного/постоянного тока) сконструирован для управления ротационным смесительным клапаном DN 15-50. Привод управляется пропорциональным сигналом и рекомендуется для установки на смесительный клапан.
Особенностью данного типа приводов является возможность передачи выходного сигнала для устройств мониторинга и управления. Также электропривод оснащён устройством, снижающим помехи входного сигнала и устройством запоминания рабочей позиции на случай временного отключения питания.
ПОДХОДЯЩИЕ СМЕСИТЕЛЬНЫЕ КЛАПАНЫ
Благодаря специальному соединению между приводом серий ARA600 и клапанами ESBE серий VRG100, VRG200 и VRB100, устройство при регулировании в целом имеет уникальную точность и устойчивость. Привод серии ARA600 также легко устанавливать на клапанах ESBE серий MG, G, F, BIV, H и HG.
- Серия VRG100
- Серия VRG200
- Серия VRG300
- Серия VRB100
- Серия MG
- Серия G
- Серия F ≤ DN50
- Серия BIV
- Серия H и HG
ПРИСОЕДИНИТЕЛЬНЫЕ КОМПЛЕКТЫ
Привод поставляется вместе с переходником для облегчения подключения к ротационному смесительному клапану ESBE. Переходники могут быть заказаны отдельно.
Переходники могут быть заказаны отдельно.
Номинальное потребление тока (A) | 8/4 |
|---|---|
Электрическое питание (В/Гц) | 24 ±10% В, 50/60 Гц |
Управляющий сигнал | 0..10 В, 2..10 В, 0..20 мА, 4..20 мА |
Время закрытия 90° (s) | 45/120 |
Крутящий момент (Нм) | 6 |
Величина вспомогательного выключателя | 6(3)А 250 В переменного тока |
Вес (кг) | 0,4 |
Степень защиты | IP41 |
Температура окружающей среды, °C | от -5 до +55 |
- Техническая спецификация
- Инструкция по монтажу
- Каталог запасных частей
Другая продукция ESBE:
- Трехходовые клапаны и комплектующие ESBE ,
- Четырехходовые клапаны ESBE ,
- Разгрузочные и защитные клапаны ESBE ,
- Электроприводы и контроллеры для клапанов ESBE ,
- Термосмесительные узлы и аксессуары ESBE ,
- Регуляторы тяги твердотопливных котлов ESBE ,
- Распределительные коллекторы для отопления (Гребенки) ESBE ,
- Насосные группы ESBE ,
- Термостатические патроны ESBE ,
- Двухходовые клапаны и комплектующие ESBE ,
- Термометры ESBE
Похожие товары
-
5,347 грн
-
5,964 грн
-
3,229 грн
-
2,774 грн
-
9,007 грн
-
6,012 грн
-
15,589 грн
-
15,589 грн
-
11,739 грн
-
7,058 грн
-
6,606 грн
-
9,791 грн
-
11,953 грн
-
15,779 грн
-
12,499 грн
-
5,347 грн
-
309 грн
-
17,133 грн
-
6,749 грн
-
12,618 грн
Смотрите также: купити твердопаливний котел львів
ЧАСТО ЗАДАВАЕМЫЕ ВОПРОСЫ ПО РОТАЦИОННЫМ ПРИВОДАМ
Привод / контроллер какого типа подходит для имеющегося клапана ESBE?
Сведения о совместимости приводов и контроллеров ESBE с клапанами ESBE см. в руководстве по выбору
в руководстве по выбору
У меня установлен клапан VRG13x. Для него требуется привод. Что порекомендуете?
Для клапанов VRG рекомендуется использовать приводы серии ARA600. Помимо типа клапана очень важно знать, какой используется управляющий сигнал. Существуют приводы с двухточечным, трехточечным или пропорциональным сигналом с электропитанием 24 В или 230 В. В большинстве контроллеров используется трехточечный управляющий сигнал с электропитанием 230 В. Настоятельно рекомендуется проверить характеристики привода в руководстве по эксплуатации котла или контроллера.
!Заявление об ограничении ответственности: привод 15sec не применяется для клапанов DN40/50
Как привод должен быть подключен к клапану ?
– Установите клапан в среднее положение. Примеры можно найти в руководстве пользователя клапана
– Установите привод на клапан, следуя инструкции пользователя привода
– Подключите привод к источнику питания после того, как удостоверитесь в верности сборки
Приводы и клапаны от ESBE всегда поставляются установленными в среднее положение.
Если привод не в среднем положении
– Переведите привод в ручной режим, затем установите привод в среднее положение
– Установите привод на клапан
– Верните привод к начальному положению и установите автоматический режим
Как можно правильно установить бывший в употреблении привод ARA6xx на смесительный клапан?
Выполните следующие действия:
- Привод: Переключитесь в ручной режим работы и поверните рукоятку в среднее положение.
- Привод: Снимите рукоятку, вытянув ее с привода. Убедитесь, что при снятии заданное положение не смещается в пункте 1 во время удаления.
- Клапан: Установите смесительный клапан в среднее положение (требуемое) и установите переходник.
- Привод: Установите привод на клапан и закрепите его болтом M5.
- Привод: Снова установите рукоятку на исполнительный механизм и, аккуратно надавливая, поверните ручку в правильное положение.
Можно ли установить привод M60 на клапан VRG?
Нет. Для обеспечения нормальной работы рекомендуется заменять привод вместе с клапаном. Найти подходящий исполнительный механизм можно в руководстве по замене в разделе «Поддержка».
Для обеспечения нормальной работы рекомендуется заменять привод вместе с клапаном. Найти подходящий исполнительный механизм можно в руководстве по замене в разделе «Поддержка».
Клапан и привод плохо работают, в чем проблема?
Удостоверьтесь в том, что клапан и привод правильно собраны вместе; больше информации относительно различных сфер применения и сборки можно найти в руководстве пользователя или на этом сайте.
Привод раскачивается взад и вперед.
Может быть несколько причин, почему привод ходит взад и вперед:
– Удостоверьтесь, что приводу подается правильный сигнал без помех
– Проверьте правильность размера клапана путем подсчета коэффициента пропускной способности на вашей системе. Подсчет можно произвести в программе ESBE Hydronic Selection, доступной на этом сайте.
Для чего можно использовать дополнительный переключатель на приводе?
Для подключения внешнего оборудования, которое вы хотели бы включать/выключать положением клапана, можно использовать микропереключатель. Например, циркуляционный насос, который надо выключить, если клапан закрыт. Или вспомогательное тепло, которое надо включить при достижении клапаном определенного положения.
Например, циркуляционный насос, который надо выключить, если клапан закрыт. Или вспомогательное тепло, которое надо включить при достижении клапаном определенного положения.
ARA600 работает, но привод и белый соединительный комплект не движутся, в чем проблема?
– Удостоверьтесь в том, что клапан и привод правильно собраны вместе; больше информации относительно различных сфер применения и сборки можно найти в руководстве пользователя или на этом сайте. Руководство пользователя доступно на странице продукта и в разделе “Загрузки”
– Удостоверьтесь, что ручка на ARA не в ручном положении. См. изображения ниже.
Что регулируется зеленым кольцом на ARA600?
Зеленым кулачковым диском включается/выключается вспомогательный переключатель.
Как изменить рабочее направление трехточечного привода ARA600?
Чтобы изменить рабочее направление, при монтаже поменяйте местами черный и коричневый провода.
Как изменить рабочее направление двухточечного привода ARA600?
С помощью перемещения перемычки на плате, например, из горизонтального в вертикальное положение.
Дополнительные сведения см. в руководстве по установке, входящем в комплект поставки, или на нашем веб-сайте.
Как изменить рабочее направление пропорционального привода ARA600?
DIP-переключатель включения/выключения 2 — ARA659
On/off on dip switch 2 – ARA659
Для дополнительной информации смотрите руководство пользователя.
Какой рабочий угол и время полного цикла выбрать?
90P поставляется с предустановленным временем полного цикла для различных рабочих углов:
- рабочий угол 90°
120 с / 90 с / 60 с
- рабочий угол 180°
240 с / 180 с / 120 с
- рабочий угол 355°
390 с / 260 с / 130 с
Какие управляющие сигналы можно установить на 90P?
Для любых настроек времени полного цикла и рабочих углов можно установить следующие управляющие сигналы:
0-10 В / 2-10 В / 0-5 В / 0-20 мА / 4-20 мА / 0-10 мА / 2-10 мА
Как настроить рабочее направление?
Рабочее направление можно установить на DIP-переключателе №5. Для движения по часовой стрелке, переключатель необходимо установить влево. Для движения против часовой стрелки, переключатель необходимо сдвинуть вправо.
Для движения по часовой стрелке, переключатель необходимо установить влево. Для движения против часовой стрелки, переключатель необходимо сдвинуть вправо.
Как настроить привод?
Чтобы задать рабочий угол, направление и время полного цикла, а также управляющий сигнал привода, смотрите в инструкции информацию обо всех DIP-переключателях и их положении.
DIP 1 и 2 – настройка рабочего угла
DIP 3 и 4 – настройка времени полного цикла
DIP 5 – настройка рабочего направления
DIP 6 и 7 – настройка управляющего сигнала
DIP 8 – настройка управляющего сигнала (напряжение или ток)
При изменении рабочего угла на DIP-переключателе, требуется дополнительная регулировка на поворотном диске, чтобы привод смог действовать в соответствии с настройками DIP-переключателя, а концевые выключатели останавливали клапан в правильном положении.
(следуйте инструкции)
Можно ли использовать ручку в любом случае и с любым рабочим углом?
Нет, ручку необходимо убрать, когда угол 180° или более.
Ротационный пропорциональный привод ESBE ARA659 12520200
Данная модель является заменой приводу ESBE 12500100.
Серия ARA600 с пропорциональным (напряжение/ток) сигналом подходит для операций смешивания совместно с клапанами VRG130 и VRB140. В этих сферах применения может быть использовано любое положение рабочего диапазона привода для достижения необходимого уровня смешивания. Привод управляется напряжением или токовым управляющим сигналом и обеспечивает более точную работу привода и клапана.
Достоинства:
- Превосходная регулировка, надежная и тихая работа.
- Возможны различные типы управляющих сигналов.
- Совершенная согласованность между приводом и клапаном ESBE.
- Доступен вспомогательный переключатель.
Технические данные
Окружающая температура:
- макс.: +55 °C
- мин.: -5 °C
Электропитание: 24 ± 10% В перем.тока/пост.тока, 50/60 Гц
Степень защиты: IP41
Класс защиты: II
Величина вспомогательного выключателя: 6(3)А 250 В переменного тока
Потребление энергии:
- Номинальное, AC: 5 Вт
- Номинальное, DC: 2.
 5 Вт
5 Вт - Допустимое, AC: ARA639, 11 ВА
- Допустимое, AC: ARA659, 8 ВА
- Допустимое, DC: ARA639, 6 ВА
- Допустимое, DC: ARA659, 4 ВА
Время действия 90° s: 45/120
Управляющий сигнал: 0..10 В, 2..10 В, 0..20 мА, 4..20 мА
Крутящий момент: 6 Нм
Производитель: ESBE AB
| Производитель | Швеция |
| Исполнение | Базовое |
| Тип | Пропорциональный |
| Режим работы | Прерывный |
| Количество фаз: | 1 |
| Класс защиты | IP41 |
| Напряжение, В | 220 |
| Частота тока, Гц | 50 / 60 |
| Диапазон рабочей температуры ВОС, °C | от -5 до +55 |
| Вес, кг | 0,4 |
История
Компания ESBE, первоначально называвшаяся Skogsfors Bruk, была основана в 1906 г. истинным предпринимателем по имени Йохан Август Скогсфорс. В возрасте всего 26 лет он запустил самое современное предприятие в южной шведской провинции Смоланд, занимавшееся производством культиваторов и сеялок – изделий, необходимых для рационализации сельского хозяйства. Более того, установив электростанцию в деревушке Рефтель, он стал первым обладателем подобной новинки.
В возрасте всего 26 лет он запустил самое современное предприятие в южной шведской провинции Смоланд, занимавшееся производством культиваторов и сеялок – изделий, необходимых для рационализации сельского хозяйства. Более того, установив электростанцию в деревушке Рефтель, он стал первым обладателем подобной новинки.
В середине 1930-х годов ассортимент продукции был расширен и стал включать в себя водяные насосы, гидрофоры и смесительные клапаны. Компания росла, и в ней начал работать Гете, сын Йохана Августа. Йохан Август решил разделить компанию на три части и передать производство смесительных клапанов Гете. Более узкоспециализированный бизнес теперь попал под управление Гете, который продолжил развитие производственных мощностей и разработку продукции. Например, в 1939 г. было запущено производство первого запатентованного 4-ходового смесительного клапана.
Вместе с развитием компании увеличивался и экспорт – смесительные клапаны ESBE скоро стали отраслевым стандартом. В 1965 г. экспорт составлял более 50% товарооборота. Кроме того, в нескольких центрально-европейских странах под словом Swedenmischer (шведские смесительные клапаны) стали подразумевать смесительные клапаны ESBE. К этому времени здоровье Гете пошатнулось, и в компании начал работать его сын Матс, представлявший третье поколение Скогсфорсов, который вскоре, в 1967 году, стал новым директором ESBE.
В 1965 г. экспорт составлял более 50% товарооборота. Кроме того, в нескольких центрально-европейских странах под словом Swedenmischer (шведские смесительные клапаны) стали подразумевать смесительные клапаны ESBE. К этому времени здоровье Гете пошатнулось, и в компании начал работать его сын Матс, представлявший третье поколение Скогсфорсов, который вскоре, в 1967 году, стал новым директором ESBE.
В середине 1970-х на рынке появились первые приводы и термостатические клапаны ESBE. Разработка продукции продолжалась, и в значительной степени она ориентировалась на потребности и пожелания потребителей, но в основе производства лежали собственные знания и идеи ESBE. В результате постоянно появлялись все более интеллектуальные решения. В качестве примера можно привести особенно удачное изделие, выпущенное в 1991 году – приводы серии M60. Благодаря этому изделию компания ESBE еще больше укрепила свои позиции на европейском рынке.
Под руководством Матса компания быстро развивалась. В конце 1990-х ее позиции в Европе существенно упрочнились благодаря открытию филиалов в Германии и Франции, а также сети специализированных торговых агентств в различных странах.
В конце 1990-х ее позиции в Европе существенно упрочнились благодаря открытию филиалов в Германии и Франции, а также сети специализированных торговых агентств в различных странах.
В 2000-х в ESBE произошло много перемен, направленных на дальнейшее усиление лидерских позиций компании. В их числе инвестиции в производственные мощности и разработка новой продукции, а также мероприятия по расширению рынка сбыта. Ярким примером является расширение сферы влияния путем открытия филиалов в Италии, Дании и Польше, а также собственных специализированных торговых представительств в Норвегии и Австрии. В то же время создано целое торговое подразделение ОЕМ-производителей комплексного оборудования, специализирующееся на международной торговле.
Продукция
РОТАЦИОННЫЕ МОТОРИЗОВАННЫЕ КЛАПАНЫ
Ротационные клапаны и приводы ESBE регулируют комфорт и тепло в домах миллионов людей. Мы предлагаем широкий ассортимент ротационных клапанов самых различных конструкций для регулировки систем отопления и охлаждения. Добавьте к этому множество различных электроприводов, просто и быстро устанавливаемых на клапанах. Обеспечивая полный контроль над устройством, они гарантируют вам надежность, экономичность и эффективность в эксплуатации на много лет вперед.
Добавьте к этому множество различных электроприводов, просто и быстро устанавливаемых на клапанах. Обеспечивая полный контроль над устройством, они гарантируют вам надежность, экономичность и эффективность в эксплуатации на много лет вперед.
КОНТРОЛЛЕРЫ
Выбор контроллеров ESBE с годами становится все разнообразнее. С появлением новых контроллеров серий CRA, CRB и CUA мы можем предложить контроль с помощью датчиков как наружной, так и внутренней температуры. И параллельно, как всегда, — регуляторы постоянной температуры. Все эти устройства объединяет одно: они обеспечивают нашим клиентам комфорт, надежность и экономичность.
ТЕРМОСТАТИЧЕСКИЕ УСТРОЙСТВА
Серии наших термостатических смесительных клапанов превратили инженеров по монтажу в героев по всей Европе. Основным требованием для обеспечения безопасности системы горячего водоснабжения является предотвращение двух важных явлений: бактерии легионеллы и ошпаривание.
ЭСБЕ | Часто задаваемые вопросы РОТАЦИОННЫЕ ПРИВОДЫ
Какой тип привода/контроллера подходит для моего клапана ESBE?
У меня есть клапан VRG13x и мне нужен привод для него.
 Чтобы вы посоветовали?
Чтобы вы посоветовали?Для клапанов VRG мы рекомендуем использовать приводы серии ARA600. Однако помимо типа клапана очень важно знать правильный управляющий сигнал. Приводы могут управляться 2-точечным, 3-точечным или пропорциональным сигналом и иметь источник питания 24 В или 230 В. Большинство контроллеров котлов используют 3-точечный управляющий сигнал с питанием 230 В, но мы настоятельно рекомендуем проверить правильные параметры привода в руководстве по эксплуатации котла или контроллера.
!Отказ от ответственности: 15-секундный привод не применим к DN40/50
Как установить привод на клапан?
– Установите клапан в среднее положение. Примеры приведены в инструкции по эксплуатации клапана
— Сборка привода на клапане в соответствии с инструкцией пользователя привода
— Сначала включите привод после того, как вы убедитесь в правильности сборки
Приводы и клапаны всегда поставляются ESBE в среднем положении.
Если привод не находится в среднем положении
– Установите привод в ручной режим и установите его в среднее положение
– Установите привод на клапан
– Верните привод в исходное положение и установите в автоматический режим
Как правильно установить бывший в употреблении привод ARA6xx на смесительный клапан?
Выполните следующие действия:
- Привод: Активируйте ручной режим работы и поверните ручку (= редуктор) в среднее положение.
- Привод: снимите ручку, вытянув ее из контроллера. При снятии следите за тем, чтобы положение, установленное в точке 1, не повернулось. Клапан
- : поверните смесительный клапан в среднее положение (желаемое) и соберите комплект переходников. Привод
- : установите привод на клапан и закрепите его болтом M5. Привод
- : установите ручку на привод и осторожно нажмите на нее, вращая ручку, чтобы найти ее правильное положение.
Можно ли установить привод M60 на клапан VRG?
Нет, это невозможно. Мы рекомендуем заменять привод вместе с клапаном для обеспечения оптимальной работы. Подходящий привод можно найти в нашем руководстве по замене, доступном в разделе «Поддержка».
Клапан и привод не работают должным образом, что не так?
Убедитесь, что клапан и привод правильно собраны вместе. Дополнительную информацию о различных применениях и способах сборки можно найти в инструкции для пользователя или на этом веб-сайте.
Привод качается вперед-назад.
Может быть несколько причин, по которым привод перемещается туда-сюда:
– Убедитесь, что сигнал на привод правильный и без каких-либо помех
– Проверьте, не слишком ли велик размер клапана, рассчитав необходимое значение KVs в вашей системе. Расчет можно выполнить в программном обеспечении ESBE Hydronic Selection, доступном на этом веб-сайте.
Для чего можно использовать вспомогательный выключатель на приводе?
Микропереключатель можно использовать для подключения внешнего оборудования, которое вы хотите запускать/останавливать по положению клапана. Примером может быть циркуляционный насос, который вы хотели бы выключить, если клапан закрыт. Другим примером может быть дополнительный обогрев, который вы хотели бы включить, когда клапан достигает определенного положения клапана.
ARA600 работает, но клапан и белый соединительный комплект не двигаются, что не так?
– Убедитесь, что клапан и привод правильно собраны вместе. Дополнительную информацию о различных применениях и способах сборки можно найти в инструкции для пользователя. Инструкция пользователя доступна на странице продукта и в папке «Область загрузки».
– Убедитесь, что ручка ARA не находится в ручном положении. Смотрите фотографии ниже.
Что регулируется зеленым кольцом на ARA600?
С помощью зеленого регулировочного кулачка вы меняете, когда вспомогательный переключатель будет активирован/деактивирован.
Как изменить рабочее направление моего 3-точечного привода ARA600 (ARA6X1; ARA6X2; ARA6X3; ARA6X4)?
Поменяв местами черный и коричневый кабель во время установки, вы можете изменить направление работы.
Как изменить рабочее направление моего привода, ARA600 2-точечный (ARA6X5; ARA6X6; ARA6X7; ARA6X8) изменен?
Сдвинув перемычки/кронштейны на печатной плате, например, из горизонтального в вертикальное.
Дополнительную информацию см. в руководстве по эксплуатации на упаковке или на нашем веб-сайте.
Как изменить рабочее направление моего пропорционального привода ARA600?
Включение/выключение двухпозиционного переключателя 6 – ARA639
Включение/выключение двухпозиционного переключателя 2 – ARA659
Более подробную информацию см. в инструкции пользователя
Можно ли регулировать рабочий диапазон на приводе серии 90?
Да, это возможно. Приводы серии 90 с 3-точечным и 2-точечным управляющим сигналом имеют настраиваемый рабочий диапазон от 30 до 180°, тип 95-270М до 270°.
Для пропорциональных версий устанавливаемый рабочий диапазон:
– между 30-90° для версии 92P
– между 30-180° для версии 92P2
– между 180-355° для версии 92P4
Обратите внимание, что эта функция не предназначена для настройки привода на неправильно отрегулированный клапан. Все приводы и контроллеры ESBE (кроме 92P4) поставляются в среднем положении. Перед монтажом привода также установите клапан в среднее положение.
Все приводы и контроллеры ESBE (кроме 92P4) поставляются в среднем положении. Перед монтажом привода также установите клапан в среднее положение.
Можно ли устанавливать привод серии 90 на клапаны серий VRG/VRB и VRH?
Да, это возможно. Используйте комплект переходников VRG801 с артикулом 16053300.
Какой рабочий угол и время работы можно выбрать?
Модель 90P поставляется с предустановленным временем работы для разных рабочих углов:
• Рабочий угол 90°
120 с / 90 с / 60 с
• Рабочий угол 180°
240 с / 180 с / 120 с
• Рабочий угол 355° 3 60 с / 23 9023
Какие управляющие сигналы можно установить на 90P?
Для всех значений времени работы и рабочих углов могут быть установлены следующие управляющие сигналы: 0-10 В / 2-10 В / 0-5 В / 0-20 мА / 4-20 мА / 0-10 мА / 2-10 мА
Как установить рабочее направление?
Направление работы можно установить с помощью DIP-переключателя №. 5. Для направления движения по часовой стрелке переключатель должен быть установлен влево. Для направления движения против часовой стрелки переключатель должен быть установлен вправо.
5. Для направления движения по часовой стрелке переключатель должен быть установлен влево. Для направления движения против часовой стрелки переключатель должен быть установлен вправо.
Как настроить привод?
Чтобы установить рабочий угол привода, направление и время работы, а также управляющий сигнал, следуйте инструкции, в которой описаны все DIP-переключатели и их положение. DIP 1 и 2 – настройка рабочего угла. DIP 3 и 4 – настройка времени работы. DIP 5 – установка рабочего направления. DIP 6 и 7 – настройка сигнала управления DIP 8 – настройка сигнала управления (напряжение или ток).
При изменении рабочего угла на DIP-переключателе требуется дополнительная регулировка вращающегося диска – это позволяет приводу двигаться в соответствии с настройками DIP-переключателя, а концевые выключатели останавливают клапан в правильном положении. (следуйте инструкции по эксплуатации).
Могу ли я использовать ручку в любом случае и под любым рабочим углом?
Нет, рукоятку следует снять, когда установлен угол или больше 180°.
MLS# 14706782 – 149 Mary Lou #a2 Drive Unit A2, Willow Park, TX 76087 – Sendero Properties, LLC
Распечатать
Виртуальный тур
Цена: 1400 долларов США
Кровати: 1
Ванны: 1
Жилая площадь кв.фут: 659
Статус: Активный
Тип свойства: Аренда жилого помещения
Год постройки: 2021
Район: Crwon Pointe Добавить Ph
дней на сайте: 291
Предоставлено Engel & Volkers Fort Worth, Рания Хадид.
Описание недвижимости
СПЕЦИАЛЬНАЯ СКИДКА на полмесяца со сроком аренды 18-24 месяца. «Заповедник в Willow Park — это активное сообщество 55+, которое представляет исключительный портрет роскошной жизни пожилых людей. Уникальное сочетание стиля и изысканности, наши жилые апартаменты отражают ваш современный стиль. Наши апартаменты 55+ Active Adult с впечатляющей отделкой и удобствами курортного качества позволяют превзойти все ваши ожидания. С таким большим количеством дел в идеальном месте вас ждет новый яркий образ жизни. Прими это. В квартирах установлены приборы из нержавеющей стали, гранитные столешницы, дизайнерские светильники, потолочные вентиляторы в спальне и гостиной, деревянные полы во всех комнатах, кроме ковра в спальнях, изготовленные на заказ деревянные шкафы, 9м потолков, книжные полки у входа.
«Заповедник в Willow Park — это активное сообщество 55+, которое представляет исключительный портрет роскошной жизни пожилых людей. Уникальное сочетание стиля и изысканности, наши жилые апартаменты отражают ваш современный стиль. Наши апартаменты 55+ Active Adult с впечатляющей отделкой и удобствами курортного качества позволяют превзойти все ваши ожидания. С таким большим количеством дел в идеальном месте вас ждет новый яркий образ жизни. Прими это. В квартирах установлены приборы из нержавеющей стали, гранитные столешницы, дизайнерские светильники, потолочные вентиляторы в спальне и гостиной, деревянные полы во всех комнатах, кроме ковра в спальнях, изготовленные на заказ деревянные шкафы, 9м потолков, книжные полки у входа.
День открытых дверей
Детали и характеристики объекта
Приборы и оборудование
- Встроенный холодильник
- Посудомоечная машина
- Утилизация
- Электрическая плита
- Электрический диапазон
- Машина, которая делает лед
- микроволновка
- подключен к льдогенератору
- Холодильник
Ванные комнаты
- 1 ванная комната
Спальни
- 1 всего
Дом
- Жилая площадь: 659
- 659 общая площадь
- Построен в 2021 году
- Детали года постройки: новое строительство – незавершенное
- Строительные материалы: камень/камень
- Структурный стиль: Квартира, Кондо/Таунхаус
Сообщество
- Клубный дом
- Общий лифт
- Общественный пул
- Общественный спринклер
- закрытый
- Беговая дорожка/велосипедная дорожка
- Прачечная
- Ограждение по периметру
Охлаждение
- Потолочные вентиляторы)
- Централ Эйр
- Электрический
Столовая
- 1 обеденная зона
Особенности инвалидности
- Установлен доступный лифт
Внешние элементы
- Балкон
- Яма огня
Забор
- Огороженный
- Ворота
Этажи
- Ковер
- Роскошная виниловая планка
Отопление
- Центральный
- Электрический
Ассоциация домовладельцев
- Тип: Нет
Элементы интерьера
- Доступно кабельное телевидение
- Декоративное освещение
- Лифт
- Доступен высокоскоростной Интернет
- Несколько лестниц
Прачечная
- Местный
Листинг
- Идентификатор листинга: 14706782
- Цена по прейскуранту: 1400 долларов США.

- Статус: Активный
- Дата доступности: 2022-01-15
- Сумма депозита: $150
- Описание срока аренды: 12+/ 24 МЕСЯЦА
- Тип аренды: 1 год плюс
- Ограничения: Другое, Ограничения для домашних животных
Местоположение
- Номер этажа: 2
- Графство или приход: Паркер
- Название подразделения: Crwon Pointe Add Ph
Лот
- 403 801 квадратный фут
- Площадь: 9
- 9,27 акра
- Размер единицы: акры
- Ландшафтный
Гостиная
- 1 жилая площадь
Энергосбережение
- Экологически чистая энергия: бытовая техника
Парковка
- 1 место в гараже
- 40 крытых мест
- Общий
- Покрытый
Правила содержания домашних животных
- Депозит: $250
- Ежемесячная плата: 20 долларов США
- Невозвращаемый сбор за размещение с домашними животными
- Разрешены домашние животные: да, ограничения по породе
Пул
- Кабана
- Пистолет
- В земле
Собственность
- Подтип: Квартира
Школы
- Район: Аледо ИСД
- Промежуточный уровень: Маканалли
- Высокий: Аледо
Система безопасности
- Пожарная тревога
- Детекторы дыма)
Налоги
- Лот: 1
- Блок: Н
- Юридическое описание: АКРЫ: 9.
 270 Участок: 1 BLK: H SUBD: CRWON POINTE
270 Участок: 1 BLK: H SUBD: CRWON POINTE
Арендатор платит
- Электричество
- Страхование
- канализация
- Вода
Коммунальные услуги
- Почтовый ящик сообщества
Windows
- оконные покрытия
Карта собственности
$1,400 – 149 Mary Lou #a2 Drive Unit A2, Willow Park, TX 76087
Просмотр улиц
Ближайшие школы
Запросить дополнительную информацию
Полное имя:
Электронная почта:
Телефон:
Сообщение:Пожалуйста, пришлите мне больше информации об этом объекте: MLS# 14706782 – $1,400 – 149Привод Мэри Лу #a2 A2, Willow Park, TX 76087.
Запланировать показ
Полное имя:
Электронная почта:
Телефон:
День и время:
Сообщение: Я хотел бы назначить встречу для показа этой собственности: MLS# 14706782 – $1,400 – 149 Mary Lou #a2 Drive Unit A2, Willow Park, TX 76087.
Похожие объекты поблизости
Последнее обновление данных листинга: 20 сентября 2022 г. 1:39PM Восточный
Поделиться на facebook
Поделиться на Twitter
Поделиться на linkedin
Поделиться на Pinterest
7 Котуитуи Драйв, Ара Хиллс, Орева, Родни | Недвижимость
Перейти к содержимому
Предыдущий Следующий
7 Kotuitui Drive, Ara Hills,
Orewa, Rodney
По договоренности
Этот потрясающий одноуровневый дом площадью 288 м² (примерно) с 6 спальнями будет построен на большом участке площадью 695 м² в популярном районе Ара Хиллс в Орева.
Гибкий дизайн обеспечивает отличное разделение, легко обслуживает семьи с маленькими детьми, подростками, расширенными семьями или получает пансион и наслаждается здоровым доходом!
• 6 больших спален с двуспальными кроватями и 3 ванные комнаты, а также гостевой туалет
• Потенциал бабушки – 2 спальни с собственной гостиной и ванной комнатой могут иметь отдельный вход
• Огромные главные спальни с ванной комнатой и гардеробной
• Идеальный высокий потолок создает ощущение пространства
• Дизайнерская кухня и умывальник с высококачественной бытовой техникой
• Большой внутренний и наружный поток, ведущий к залитой солнцем открытой площадке на северной стороне.
• Гараж на две машины с внутренним доступом
Ara Hills превосходно расположен прямо у развязки Орева на государственной автомагистрали 1, что дает доступ к поездке на север или юг, являясь полной развязкой и всего в 34 км (приблизительно) от центрального делового района Окленда. Всего в 3 км от пляжа Орева и деревни, где предлагаются все удобства, включая спортивные клубы, кафе и рестораны, искусство и мероприятия, бизнес-услуги и многое другое. Сильвердейл находится в 5 минутах езды от торгового центра Silverdale Mall, торгового центра, кафе, бассейна и тренажерного зала Northern Arena, Snow Planet, а также ряда крупных розничных магазинов, таких как Bunnings, Plant Barn, Farmers, Briscoes…..
Всего в 3 км от пляжа Орева и деревни, где предлагаются все удобства, включая спортивные клубы, кафе и рестораны, искусство и мероприятия, бизнес-услуги и многое другое. Сильвердейл находится в 5 минутах езды от торгового центра Silverdale Mall, торгового центра, кафе, бассейна и тренажерного зала Northern Arena, Snow Planet, а также ряда крупных розничных магазинов, таких как Bunnings, Plant Barn, Farmers, Briscoes…..
Не медлите, свяжитесь с нами сейчас для получения более подробной информации!
Хотели бы вы приехать и посмотреть эту недвижимость?
ЗАПРОСИТЬ ПРОСМОТР
Характеристики недвижимости
| Идентификатор недвижимости: | ПНх40149 |
| Земля: | 695 м 2 |
| Спальни: | 6 |
| Ванные комнаты: | 3 |
| Здание: | 288м 2 |
| Автор списка: | Лицензия Access Real Estate Ltd (REAA 2008) |
| Поделиться: |
Свяжитесь с нами
Фиона Гао
Лицензиат Продавец и управляющий недвижимостью
Рэй Уайт Пайнхилл
+64 21 0292 показать
+64 (9) 477 показать
ЗАДАЙТЕ ВОПРОС
Джимми Ву
Агент Лицензиата / Принципал
Рэй Уайт Пайнхилл
+64 21 659показать
+64 (9) 947 показать
ЗАДАЙТЕ ВОПРОС
Имя *
Фамилия
Номер телефона
Электронная почта *
Запрос *
Имя *
Фамилия
Номер телефона
Электронная почта *
Дата и время просмотра
Другие имена участников
Ваш текущий адрес проживания *
Ваш текущий статус * У меня есть финансирование Я еще не организовал финансирование Другой
Чтобы обеспечить здоровье и безопасность всех сторон, мы просим вас ознакомиться и принять наши условия просмотра недвижимости.
Условия просмотра недвижимости:
- Те, кто плохо себя чувствует, самоизолируются или находятся в домашнем хозяйстве, могут не присутствовать на просмотре.
- Рекомендуется физическое дистанцирование и соблюдение правил гигиены.
- Рекомендуется ношение маски для лица.
Я/мы прочитал(а) и согласен(на) со всеми положениями и условиями посещения осмотра недвижимости, изложенными выше.
Механизмы устойчивости к 5-аза-2′-дезоксицитидину в линиях раковых клеток человека | Кровь
Пропустить пункт назначения Nav
МИЕЛОИДНАЯ НЕОПЛАЗИЯ| 15 января 2009 г.
Тайчунь Цинь,
Ярослав Елинек,
Джиали Си,
Цзинмин Шу,
Жан-Пьер Ж. Исса
Исса
Кровь (2009) 113 (3): 659–667.
https://doi.org/10.1182/blood-2008-02-140038
История статьи
Подано:
19 февраля 2008 г.
Принято:
27 августа 2008 г.
Первая редакция:
17 октября 2008 г. Разделенный экран
- Твиттер
Запросить разрешения
Иконка Цитировать Цитировать
Цитирование
Тайчунь Цинь, Ярослав Елинек, Джиали Си, Цзинмин Шу, Жан-Пьер Ж. Исса; Механизмы устойчивости к 5-аза-2′-дезоксицитидину в линиях раковых клеток человека. Кровь 2009; 113 (3): 659–667. дои: https://doi.org/10.1182/blood-2008-02-140038
Исса; Механизмы устойчивости к 5-аза-2′-дезоксицитидину в линиях раковых клеток человека. Кровь 2009; 113 (3): 659–667. дои: https://doi.org/10.1182/blood-2008-02-140038
Скачать файл цитаты:
- Рис (Зотеро)
- Менеджер ссылок
- EasyBib
- Подставки для книг
- Менделей
- Бумаги
- Конечная примечание
- РефВоркс
- Бибтекс
5-аза-2′-дезоксицитидин (DAC) одобрен для лечения миелодиспластических синдромов, но резистентность к этому агенту является обычным явлением. В поисках механизмов резистентности измеряли полумаксимальную (50%) ингибирующую концентрацию (IC 50 ) DAC и обнаружили, что он отличается в 1000 раз среди группы линий раковых клеток. IC 50 коррелировало с дозами DAC, которые вызывали наибольшее гипометилирование длинных вкрапленных ядерных элементов (LINE; R = 0,94, P <0,001), но не с метилированием LINE или ДНК-метилтрансферазой 1 (DNMT1).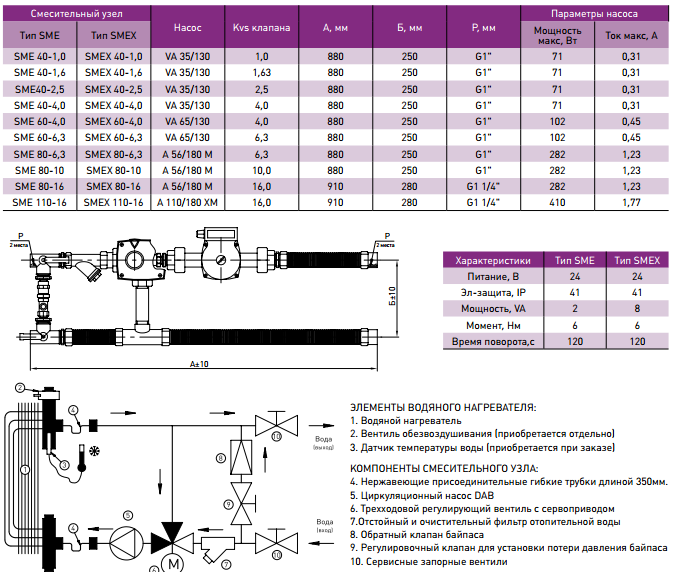 3a и 3b экспрессия на исходном уровне. Чувствительность к DAC показала низкую корреляцию (R = 0,44, P = 0,11) с чувствительностью к 5-азацитидину (AZA), но хорошую корреляцию с цитарабином (Ara-C; R = 0,89)., P < 0,001). 5 клеточных линий, наиболее устойчивых к DAC, имели сочетание низкого уровня транспортеров dCK, hENT1 и 2 и высокого уровня цитозиндезаминазы. В клоне HL60 резистентность к DAC могла быстро индуцироваться воздействием лекарственного средства и была связана с переходом от гетерозиготной к гомозиготной мутации DCK . Трансфекция дикого типа DCK восстанавливала чувствительность DAC. DAC индуцирует разрывы ДНК, о чем свидетельствует фосфорилирование h3AX и увеличение скорости гомологичной рекомбинации в 7–10 раз. Эти результаты позволяют предположить, что устойчивость in vitro к DAC можно объяснить недостаточным включением в ДНК.
3a и 3b экспрессия на исходном уровне. Чувствительность к DAC показала низкую корреляцию (R = 0,44, P = 0,11) с чувствительностью к 5-азацитидину (AZA), но хорошую корреляцию с цитарабином (Ara-C; R = 0,89)., P < 0,001). 5 клеточных линий, наиболее устойчивых к DAC, имели сочетание низкого уровня транспортеров dCK, hENT1 и 2 и высокого уровня цитозиндезаминазы. В клоне HL60 резистентность к DAC могла быстро индуцироваться воздействием лекарственного средства и была связана с переходом от гетерозиготной к гомозиготной мутации DCK . Трансфекция дикого типа DCK восстанавливала чувствительность DAC. DAC индуцирует разрывы ДНК, о чем свидетельствует фосфорилирование h3AX и увеличение скорости гомологичной рекомбинации в 7–10 раз. Эти результаты позволяют предположить, что устойчивость in vitro к DAC можно объяснить недостаточным включением в ДНК.
Темы:
клеточных линий, цитарабин, дезоксицитидин, дезоксицитидинкиназа, ДНК, трансфекция, опухолевые клетки, злокачественные, клонированные клетки, днк (цитозин-5-)-метилтрансфераза 1, метилирование
Эпигенетические изменения все чаще признаются движущей силой лейкемии человека. 1-3 Например, аномальное метилирование со временем накапливается в различных участках генома и способствует онкогенезу за счет увеличения нестабильности генома или подавления генов-супрессоров опухолей. Замалчивание генов-супрессоров опухолей тесно связано с гиперметилированием; метилированные гены-супрессоры опухолей могут быть реактивированы ингибиторами ДНК-метилтрансферазы (DNMT). Эти наблюдения привели к возрождению интереса к ингибиторам метилирования ДНК в качестве противоопухолевых средств в клинических испытаниях. 4 Прототипы ингибиторов DNMT 5-аза-2′-дезоксицитидин (DAC) и 5-азацитидин (AZA) недавно были одобрены Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США в качестве противоопухолевых средств для лечения миелодиспластического синдрома. 5,6 Одна потенциальная проблема с обоими агентами заключается в том, что во время лечения может развиться резистентность. Оказавшись внутри клетки, DAC фосфорилируется дезоксицитидинкиназой (dCK) в монофосфорилированное производное 5-аза-dCMP.
1-3 Например, аномальное метилирование со временем накапливается в различных участках генома и способствует онкогенезу за счет увеличения нестабильности генома или подавления генов-супрессоров опухолей. Замалчивание генов-супрессоров опухолей тесно связано с гиперметилированием; метилированные гены-супрессоры опухолей могут быть реактивированы ингибиторами ДНК-метилтрансферазы (DNMT). Эти наблюдения привели к возрождению интереса к ингибиторам метилирования ДНК в качестве противоопухолевых средств в клинических испытаниях. 4 Прототипы ингибиторов DNMT 5-аза-2′-дезоксицитидин (DAC) и 5-азацитидин (AZA) недавно были одобрены Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США в качестве противоопухолевых средств для лечения миелодиспластического синдрома. 5,6 Одна потенциальная проблема с обоими агентами заключается в том, что во время лечения может развиться резистентность. Оказавшись внутри клетки, DAC фосфорилируется дезоксицитидинкиназой (dCK) в монофосфорилированное производное 5-аза-dCMP. Впоследствии 5-аза-dCMP фосфорилируется до его активной формы, 5-aza-dCTP, которая включается в ДНК, где вызывает деметилирование. Метаболиты DAC также могут быть субстратами для ферментов катаболизма, таких как цитидиндезаминаза (CDA), которые катализируют инактивацию цитидина и дезоксицитидина до уридина и дезоксиуридина, тем самым уменьшая количество 5-аза-dCTP, которое может образоваться. Аналог нуклеозида Ara-C также зависит от dCK в качестве начальной стадии, ограничивающей скорость включения. Однако AZA не нуждается в dCK. Его включение зависит от уридин-цитидинкиназы (UCK).
Впоследствии 5-аза-dCMP фосфорилируется до его активной формы, 5-aza-dCTP, которая включается в ДНК, где вызывает деметилирование. Метаболиты DAC также могут быть субстратами для ферментов катаболизма, таких как цитидиндезаминаза (CDA), которые катализируют инактивацию цитидина и дезоксицитидина до уридина и дезоксиуридина, тем самым уменьшая количество 5-аза-dCTP, которое может образоваться. Аналог нуклеозида Ara-C также зависит от dCK в качестве начальной стадии, ограничивающей скорость включения. Однако AZA не нуждается в dCK. Его включение зависит от уридин-цитидинкиназы (UCK).
Дефицит dCK является основным известным механизмом резистентности к цитидиновым NA in vitro и, как сообщается, связан с резистентностью in vivo у некоторых пациентов. 7,8 Например, индуцированная in vitro резистентность к аналогам дезоксицитидина цитарабину (Ara-C) и DAC в крысиной модели острого миелоидного лейкоза была опосредована мутациями в гене DCK . 9 Однако мало информации о происхождении этого вида сопротивления. Резистентность к лечению противоопухолевыми препаратами является результатом множества факторов, включая спонтанную генетическую нестабильность в опухолях и механизмы индукции лекарств, которые, вероятно, играют важную роль в приобретенной устойчивости к противоопухолевым препаратам. 10-16 Дефекты метилирования ДНК могут способствовать нестабильности генома, что приводит к повышенной частоте мутаций. 17-20 Таким образом, мы предположили, что как спонтанная генетическая нестабильность, так и генетическая нестабильность, индуцированная DAC, способствуют возникновению устойчивости к DAC в линиях раковых клеток человека и что устойчивость к DAC обусловлена недостаточным внутриклеточным трифосфатом DAC. Чтобы проверить эти гипотезы, мы исследовали in vitro модели естественной устойчивости к DAC в различных линиях раковых клеток и дополнительно исследовали механизмы и происхождение устойчивости в клеточной линии миелоидного лейкоза HL60.
Резистентность к лечению противоопухолевыми препаратами является результатом множества факторов, включая спонтанную генетическую нестабильность в опухолях и механизмы индукции лекарств, которые, вероятно, играют важную роль в приобретенной устойчивости к противоопухолевым препаратам. 10-16 Дефекты метилирования ДНК могут способствовать нестабильности генома, что приводит к повышенной частоте мутаций. 17-20 Таким образом, мы предположили, что как спонтанная генетическая нестабильность, так и генетическая нестабильность, индуцированная DAC, способствуют возникновению устойчивости к DAC в линиях раковых клеток человека и что устойчивость к DAC обусловлена недостаточным внутриклеточным трифосфатом DAC. Чтобы проверить эти гипотезы, мы исследовали in vitro модели естественной устойчивости к DAC в различных линиях раковых клеток и дополнительно исследовали механизмы и происхождение устойчивости в клеточной линии миелоидного лейкоза HL60.
Культура клеток и протоколы лечения
Линии клеток лейкемии и лимфомы человека HL60, ML-1, HEL, Raji, Jurkat, TF-1, U937, K562 и MOLT4; клеточные линии рака предстательной железы PC3 и DU145; клеточные линии рака толстой кишки RKO и SW48; и линия клеток рака молочной железы Cama-1 были получены от ATCC (Манассас, Вирджиния). Клетки выращивали в RPMI 1640 плюс 10% термоинактивированной фетальной телячьей сыворотки (FCS) в пластиковых планшетах для тканевых культур во влажной атмосфере, содержащей 5% CO 2 , при 37°C. Для анализа ингибирования роста клетки помещали с плотностью 2,5 × 10 9 .0692 5 /мл в 5 мл среды за 24 часа до лечения. К среде добавляли градуированные концентрации DAC. Для измерения половины максимальной (50%) ингибирующей концентрации (IC 50 ) свежий DAC добавляли каждые 24 часа без смены среды. Дозы, ингибирующие пролиферацию на 50% (IC 50 ), анализировали методом медианного эффекта с помощью программного обеспечения CalcuSyn (Biosoft, Cambridge, United Kingdom). 21 Долю живых клеток в обработанных планшетах измеряли методом исключения трипанового синего.
Клетки выращивали в RPMI 1640 плюс 10% термоинактивированной фетальной телячьей сыворотки (FCS) в пластиковых планшетах для тканевых культур во влажной атмосфере, содержащей 5% CO 2 , при 37°C. Для анализа ингибирования роста клетки помещали с плотностью 2,5 × 10 9 .0692 5 /мл в 5 мл среды за 24 часа до лечения. К среде добавляли градуированные концентрации DAC. Для измерения половины максимальной (50%) ингибирующей концентрации (IC 50 ) свежий DAC добавляли каждые 24 часа без смены среды. Дозы, ингибирующие пролиферацию на 50% (IC 50 ), анализировали методом медианного эффекта с помощью программного обеспечения CalcuSyn (Biosoft, Cambridge, United Kingdom). 21 Долю живых клеток в обработанных планшетах измеряли методом исключения трипанового синего.
Пиросеквенирование
Мы использовали ДНК-повторяющийся длинный вкрапленный ядерный элемент (LINE) в качестве маркера и анализ метилирования на основе пиросеквенирования для изучения глобального метилирования геномной ДНК, как описано ранее. 5 Геномную ДНК получали из клеток и проводили бисульфитную конверсию геномной ДНК. LINE амплифицировали с помощью ПЦР с использованием прямого праймера 5′-TTTTTTGAGTTAGGTGTGGG-3′ и 5′-биотинилированного обратного праймера 5′-TCTCACTAAAAAATACCAAACAA-3′. После ПЦР биотинилированная обратная цепь была захвачена на гранулах стрептавидин-сефарозы (Amersham Biosciences, Уппсала, Швеция) и отожжена с праймером для секвенирования 5′-GGGTGGGAGTGAT-3′. Для измерения потери гетерозиготности (LOH) из-за гетерозиготной мутации в экзоне 3 из DCK , мы использовали основанный на пиросеквенировании анализ точечной мутации в нуклеотиде 454 мРНК DCK (NM_000788) и соседнего однонуклеотидного полиморфизма (SNP) в нуклеотиде 459 (rs11544786). Экзон 3 DCK амплифицировали из геномной ДНК и кДНК с помощью ПЦР с использованием прямого праймера 5′-GGTGGGAATGTTCTTCAGA-3′ и обратного праймера 5′-AGCCATTTATACATACCTGTCAC-3′. Продукт ПЦР использовали в качестве матрицы, а затем амплифицировали с прямым праймером 5′-GGTGGGAATGTTCTTCAGA-3′, обратным праймером 5′-GGGACACCGCTGATCGTTTATTTAGCCATTTATACATACCTGTCAC-3′ и 5′-биотинилированным универсальным праймером 5′-GGGACACCGCTGATCGTTTA-3′.
5 Геномную ДНК получали из клеток и проводили бисульфитную конверсию геномной ДНК. LINE амплифицировали с помощью ПЦР с использованием прямого праймера 5′-TTTTTTGAGTTAGGTGTGGG-3′ и 5′-биотинилированного обратного праймера 5′-TCTCACTAAAAAATACCAAACAA-3′. После ПЦР биотинилированная обратная цепь была захвачена на гранулах стрептавидин-сефарозы (Amersham Biosciences, Уппсала, Швеция) и отожжена с праймером для секвенирования 5′-GGGTGGGAGTGAT-3′. Для измерения потери гетерозиготности (LOH) из-за гетерозиготной мутации в экзоне 3 из DCK , мы использовали основанный на пиросеквенировании анализ точечной мутации в нуклеотиде 454 мРНК DCK (NM_000788) и соседнего однонуклеотидного полиморфизма (SNP) в нуклеотиде 459 (rs11544786). Экзон 3 DCK амплифицировали из геномной ДНК и кДНК с помощью ПЦР с использованием прямого праймера 5′-GGTGGGAATGTTCTTCAGA-3′ и обратного праймера 5′-AGCCATTTATACATACCTGTCAC-3′. Продукт ПЦР использовали в качестве матрицы, а затем амплифицировали с прямым праймером 5′-GGTGGGAATGTTCTTCAGA-3′, обратным праймером 5′-GGGACACCGCTGATCGTTTATTTAGCCATTTATACATACCTGTCAC-3′ и 5′-биотинилированным универсальным праймером 5′-GGGACACCGCTGATCGTTTA-3′. Биотинилированную цепь отжигали с праймером для секвенирования 5′-TGGTCTTTTACCTTCCA-3′. Пиросеквенирование проводили с использованием PSQ HS 9.6 Реагенты Gold SNP и устройство для пиросеквенирования PSQ HS 96 (Biotage, Уппсала, Швеция).
Биотинилированную цепь отжигали с праймером для секвенирования 5′-TGGTCTTTTACCTTCCA-3′. Пиросеквенирование проводили с использованием PSQ HS 9.6 Реагенты Gold SNP и устройство для пиросеквенирования PSQ HS 96 (Biotage, Уппсала, Швеция).
Измерение частоты резистентности
Клетки HL60 высевали на 6-луночные планшеты по 10 5 клеток на лунку в среде Iscove с добавлением 1% метилцеллюлозы, 10% эмбриональной бычьей сыворотки, 2 мМ l-глютамина, 100 МЕ/ мл пенициллина, 100 мкг/мл стрептомицина и 10 мкМ DAC для отбора резистентных колоний. Колонии подсчитывали через 14 дней. Частоту резистентности оценивали как процент резистентных колоний, деленный на эффективность посева в среде без DAC.
Измерение продукции трифосфата Ara-C
Производство трифосфата Ara-C (Ara-CTP) измеряли, как описано ранее. 22 Вкратце, клетки HL60 обрабатывали 10 мМ [ 3 H] Ara-CTP в течение 4 часов и центрифугировали при 1000× g в течение 10 минут. Осадки промывали и экстрагировали трихлоруксусной кислотой для удаления белков. Кислый экстракт нейтрализовали, а водный слой использовали для разделения с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ). Ara-CTP определяли количественно при 262 нм путем электронного интегрирования со ссылкой на внешние стандарты.
Осадки промывали и экстрагировали трихлоруксусной кислотой для удаления белков. Кислый экстракт нейтрализовали, а водный слой использовали для разделения с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ). Ara-CTP определяли количественно при 262 нм путем электронного интегрирования со ссылкой на внешние стандарты.
Измерение активности dCK
Активность dCK измеряли, как описано ранее. 23 Клетки HL60 центрифугировали при 1000 g в течение 10 минут, ресуспендировали в буфере А (50 мМ фосфата калия, рН 7,5 и 10 мМ 2-меркаптоэтанол) и обрабатывали ультразвуком. Суспензию центрифугировали при 145 000 g в течение 20 минут. Супернатант использовали в виде сырого экстракта после диализа против 100 объемов буфера А. К 100 мкл реакционной смеси (50 мМ Трис-HCl, рН 7,8; 5 мМ MgCl 9 ) добавляли 0,1 мг общего белка.0919 2 , 8 мМ UTP и 25 мкМ [ 3 H]Ara-C [4 Ci/ммоль]), а затем инкубировали смесь при 37°C. С 10-минутными интервалами образцы по 20 мкл отбирали (4 раза) и наносили на диски, покрытые диэтиламиноэтилом (DEAE) (DE-81; Whatman, Maidstone, United Kingdom), которые затем промывали 3 раза в течение 10–15 минут. в 1 мМ формиате аммония, дважды с деионизированным H 2 O 2 и один раз с 95% этанолом, а затем сушили. Радиоактивность подсчитывали в 10 мл Aquasol. Удельную активность выражали в виде распадов в минуту (DPM) на миллиграмм белка.
С 10-минутными интервалами образцы по 20 мкл отбирали (4 раза) и наносили на диски, покрытые диэтиламиноэтилом (DEAE) (DE-81; Whatman, Maidstone, United Kingdom), которые затем промывали 3 раза в течение 10–15 минут. в 1 мМ формиате аммония, дважды с деионизированным H 2 O 2 и один раз с 95% этанолом, а затем сушили. Радиоактивность подсчитывали в 10 мл Aquasol. Удельную активность выражали в виде распадов в минуту (DPM) на миллиграмм белка.
Вестерн-блот-анализ
Для вестерн-блоттинга клеточные лизаты смешивали с таким же объемом 2× буфера для образцов Лэммли (Bio-Rad Laboratories, Hercules, CA), кипятили и наносили на 10% полиакриламидные гели, содержащие додецилсульфат натрия. . Белки переносили на поливинилиденфторидные мембраны. Мы использовали кроличье антитело против фосфогистона h3AX (Sigma-Aldrich, Сент-Луис, Миссури) и антитела DNMT1, 3a и 3b (Abcam, Кембридж, Массачусетс) и кроличье поликлональное антитело к dCK (щедрый подарок доктора Кеслера Гергели из Университета Земмельвайса). , Будапешт, Венгрия).
, Будапешт, Венгрия).
Мы клонировали полную кодирующую область DCK дикого типа в сайты рестрикционных ферментов Bgl II и Eco R1 с использованием вектора pEGFP-N1 (Clontech, Mountain View, КА). Полноразмерную кДНК DCK амплифицировали с помощью полимеразной цепной реакции с обратной транскрипцией (ОТ-ПЦР) с использованием прямого праймера 5′-TATCTCAGATCTTTGCCGGACGAGCTCTG-3′ и обратного праймера 5′-ATTGAATTCTGGAACCATTTGGCTGCCTG-3′. Затем мы трансфицировали этот вектор в dCK-дефицитные клетки HL60 с использованием реагента Lipofectamine 2000 (Invitrogen, Carlsbad, CA) в соответствии с инструкциями производителя. Антибиотик G418 в концентрации 0,5 мг/мл (American Bioanalytical, Natick, MA) добавляли в среду для культивирования клеток для отбора клеток со стабильной интеграцией вектора, содержащего ДСК . Имитация трансфекции пустым вектором pEGFP-N1 использовалась в качестве контроля.![]()
Измерение гомологичной рекомбинационной репарации
Чтобы определить, может ли DAC индуцировать гомологичную рекомбинационную репарацию (HRR), мы трансфицировали векторы pLNCX-GZ, pHit60 и pVSV-G в клетки почки эмбриона человека 293T, как описано ранее. 24 Через три дня после трансфекции собирали среду, содержащую ретровирус. Клетки HL60 инкубировали с разведениями ретровирусного вектора в RPMI 1640. В среду для культивирования клеток добавляли антибиотик G418 (0,5 мг/мл) для отбора клеток со стабильной интеграцией LNCX-GZ. Затем мы обрабатывали клетки 0,02, 0,2, 2 или 20 мкМ DAC в течение 4 дней и поддерживали их в среде без DAC, содержащей G418, в течение 10 дней. Всего 3 × 10 9Клетки 0692 5 добавляли к 3 мл метилцеллюлозы с G418 плюс антибиотик зеоцин 200 мкг/мл (Invitrogen, Карлсбад, Калифорния) для селекции. Клетки осторожно перемешивали на вортексе, чтобы они равномерно суспендировались, а затем высевали на 6-луночные планшеты. Колонии подсчитывали через 14 дней. Частоту рекомбинации оценивали путем деления количества колоний, устойчивых к G418 и зеоцину, на общее количество загруженных и снова деления на эффективность посева. Праймеры, фланкирующие 2 кассеты зеленого флуоресцентного белка (GFP)-зеоцин, использовали для идентификации ожидаемых событий рекомбинации. Прямой праймер представлял собой 5′-GCTAGCTTGCCAAACCTACAG-3′, а обратный праймер представлял собой 5′-GTGAACCGTCAGATCCGCTAG-3′. Для нерекомбинированного вектора ПЦР-фрагмент составлял 2,2 т.п.о., тогда как фрагмент размером 1,1 т.п.о. был получен рекомбинированным вектором.
Колонии подсчитывали через 14 дней. Частоту рекомбинации оценивали путем деления количества колоний, устойчивых к G418 и зеоцину, на общее количество загруженных и снова деления на эффективность посева. Праймеры, фланкирующие 2 кассеты зеленого флуоресцентного белка (GFP)-зеоцин, использовали для идентификации ожидаемых событий рекомбинации. Прямой праймер представлял собой 5′-GCTAGCTTGCCAAACCTACAG-3′, а обратный праймер представлял собой 5′-GTGAACCGTCAGATCCGCTAG-3′. Для нерекомбинированного вектора ПЦР-фрагмент составлял 2,2 т.п.о., тогда как фрагмент размером 1,1 т.п.о. был получен рекомбинированным вектором.
В этом исследовании мы исследовали механизмы внутренней устойчивости к DAC в группе линий раковых клеток и приобретенной устойчивости в клеточной линии HL60. Во-первых, мы выбрали несколько клеточных линий лейкемии, поскольку DAC наиболее активен при лейкемии, а также включили клеточные линии рака толстой кишки, молочной железы и простаты для представления солидных опухолей. Мы измерили IC 50 DAC, Ara-C и AZA в различных клеточных линиях. IC 50 DAC составляла менее 0,05 мкМ в TF-1, U937, Raji и HEL; от 0,05 до 0,4 мкМ в ML-1, HL-60, K562, SW48 и Cama-1; и более 2 мкМ в Jurkat, MOLT4, PC3, RKO и DU145, которые были определены как устойчивые клеточные линии (таблица 1). ИС 50 DAC коррелировал с таковым для Ara-C (R = 0,89, P < 0,001), но незначительно с чувствительностью к AZA (R = 0,44, P = 0,11; рисунок 1A).
Мы измерили IC 50 DAC, Ara-C и AZA в различных клеточных линиях. IC 50 DAC составляла менее 0,05 мкМ в TF-1, U937, Raji и HEL; от 0,05 до 0,4 мкМ в ML-1, HL-60, K562, SW48 и Cama-1; и более 2 мкМ в Jurkat, MOLT4, PC3, RKO и DU145, которые были определены как устойчивые клеточные линии (таблица 1). ИС 50 DAC коррелировал с таковым для Ara-C (R = 0,89, P < 0,001), но незначительно с чувствительностью к AZA (R = 0,44, P = 0,11; рисунок 1A).
Таблица 1
IC 50 DAC, AZA и Ara-C в линиях раковых клеток человека
| Линия клеток . | Ткань . | ИК 50 , нМ . | LINE плотность метилирования % . | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ЦАП . | АЗА . | Ара-С
. | |||||||||||||||||||||||||||||
| TF-1 | Leukemia | 10 ± 2.1 | 76 ± 22 | 0.45 ± 0.15 | 28 | ||||||||||||||||||||||||||
| U93 7 | Lymphoma | 10 ± 1.8 | 281 ± 34 | 0,99 ± 0,24 | 62 | ||||||||||||||||||||||||||
| HEL | Лейкемия | 40 ± 2,5 | 65 ± 12 | 0.65 ± 0.12 | 54 | ||||||||||||||||||||||||||
| Raji | Lymphoma | 54 ± 21 | 2332 ± 264 | 1.6 ± 0.32 | 75 | ||||||||||||||||||||||||||
| Cama-1 | Breast | 65 ± 7,0 | 280 ± 33 | 16 ± 1,5 | 30 | ||||||||||||||||||||||||||
| SW48 | Colon | 100 ± | 7 10. | 100 ± |  “>7 10. “>7 10. | 100 ± | 7 10. 101097. | 100 ± | 7 | 100 ± | 100 ± | 100 ± | .0687 | ||||||||||||||||||
| ML-1 | Leukemia | 98 ± 27 | 612 ± 55 | 1.7 ± 0.19 | 58 | ||||||||||||||||||||||||||
| HL60 | Leukemia | 200 ± 54 | 96 ± 37 | 3.7 ± 0.61 | 75 | ||||||||||||||||||||||||||
| K562 | Leukemia | 400 ± 34 | 102 ± 9.8 | 1.8 ± 0.25 | 12 | ||||||||||||||||||||||||||
| MOLT4 | Leukemia | 1802 ± 125 | 680 ± 112 | 10 ± 0.31 | 80 | ||||||||||||||||||||||||||
| Jurkat | Leukemia | 2000 ± 209 |  “> 65 ± 15 “> 65 ± 15 | 17.2 ± 0.91 | 75 | ||||||||||||||||||||||||||
| PC3 | Prostate | 7501 ± 550 | 5600 ± 259 | 210 ± 22 | 44 | ||||||||||||||||||||||||||
| RKO | Colon | 9909 ± 980 | 2100 ± 147 | 20 ± 24 | 54 | ||||||||||||||||||||||||||
| DU145 | Простата | 10 000 ± 660 | 2822 ± 330 | 300 ± 26 | 66 | 66 | 66 | . . | Ткань . | ИК 50 , нМ . | LINE плотность метилирования % . | ||||||||||||||||||||
| ЦАП . | АЗА . | Ара-С
. | |||||||||||||||||||||||||||||
| TF-1 | Leukemia | 10 ± 2.1 | 76 ± 22 | 0.45 ± 0.15 | 28 | ||||||||||||||||||||||||||
| U93 7 | Lymphoma | 10 ± 1.8 | 281 ± 34 | 0.99 ± 0.24 | 62 | ||||||||||||||||||||||||||
| HEL | Leukemia | 40 ± 2.5 | 65 ± 12 | 0.65 ± 0.12 | 54 | ||||||||||||||||||||||||||
| Raji | Lymphoma | 54 ± 21 | 2332 ± 264 | 1.6 ± 0.32 | 75 | ||||||||||||||||||||||||||
| Cama-1 | Breast | 65 ± 7.0 | 280 ± 33 | 16 ± 1.5 | 30 | ||||||||||||||||||||||||||
| SW48 | Colon | 100 ± 9.1 | 101 ± 12 |  “> 2.1 ± 0.21 “> 2.1 ± 0.21 | 62 | ||||||||||||||||||||||||||
| ML-1 | Leukemia | 98 ± 27 | 612 ± 55 | 1.7 ± 0.19 | 58 | ||||||||||||||||||||||||||
| HL60 | Leukemia | 200 ± 54 | 96 ± 37 | 3.7 ± 0.61 | 75 | ||||||||||||||||||||||||||
| K562 | Leukemia | 400 ± 34 | 102 ± 9.8 | 1.8 ± 0,25 | 12 | ||||||||||||||||||||||||||
| MOLT4 | Лейкемия | 1802 ± 125 | 680 ± 112 | 10 ± 0,31 | 10 ± 0,314| 10 ± 0,31 | .0684 | 2000 ± 209 | 65 ± 15 | 17.2 ± 0.91 | 75 | PC3 | Prostate |  “> 7501 ± 550 “> 7501 ± 550 5600 ± 259 | 210 ± 22 | 44 | RKO | Colon | 9909 ± 980 | 2100 ± 147 | 20 ± 24 | 54 | DU145 | PROSTATE | DU145 | .0684 | 66 | | ||||
Значения представлены как среднее плюс или минус SEM из 3 независимых экспериментов.
Увеличить
Рисунок 1
Увеличить Загрузить PPT
Дозозависимая индукция гипометилирования DAC в различных клеточных линиях . (A) IC 50 DAC, AZA и Ara-C в линиях раковых клеток человека. Мы измерили IC 50 DAC, AZA и Ara-C на панели линий раковых клеток человека и сопоставили IC 50 DAC по сравнению с IC 50 Ara-C, IC 50 DAC по сравнению с IC 50 AZA соответственно. (B) Дозозависимая индукция гипометилирования DAC в различных клеточных линиях. После обработки DAC в течение 4 дней клетки собирали и выделяли ДНК. Метилирование LINE измеряли с помощью анализа бисульфитного пиросеквенирования. В каждой клеточной линии, кроме наиболее устойчивых клеток (нижний график), кривая зависимости от дозы имела U-образную форму. (B) Отсутствие корреляции IC 50 DAC с метилированием LINE на исходном уровне (R = 0,05, P = 0,97). (C) Корреляция между IC 50 DAC с дозами DAC, необходимыми для максимального гипометилирования LINE (R = 0,94, P <0,001).
После обработки DAC в течение 4 дней клетки собирали и выделяли ДНК. Метилирование LINE измеряли с помощью анализа бисульфитного пиросеквенирования. В каждой клеточной линии, кроме наиболее устойчивых клеток (нижний график), кривая зависимости от дозы имела U-образную форму. (B) Отсутствие корреляции IC 50 DAC с метилированием LINE на исходном уровне (R = 0,05, P = 0,97). (C) Корреляция между IC 50 DAC с дозами DAC, необходимыми для максимального гипометилирования LINE (R = 0,94, P <0,001).
Рисунок 1
Просмотреть в большом формате Скачать PPT
Дозозависимая индукция гипометилирования с помощью DAC в различных клеточных линиях . (A) IC 50 DAC, AZA и Ara-C в линиях раковых клеток человека. Мы измерили IC 50 DAC, AZA и Ara-C на панели линий раковых клеток человека и корреляция IC 50 DAC по сравнению с IC 50 Ara-C, IC 50 DAC по сравнению с IC 50 АЗА соответственно. (B) Дозозависимая индукция гипометилирования DAC в различных клеточных линиях. После обработки DAC в течение 4 дней клетки собирали и выделяли ДНК. Метилирование LINE измеряли с помощью анализа бисульфитного пиросеквенирования. В каждой клеточной линии, кроме наиболее устойчивых клеток (нижний график), кривая зависимости от дозы имела U-образную форму. (B) Отсутствие корреляции IC 50 DAC с метилированием LINE на исходном уровне (R = 0,05, P = 0,97). (C) Корреляция между IC 50 DAC с дозами DAC, необходимыми для максимального гипометилирования LINE (R = 0,94, P <0,001).
(B) Дозозависимая индукция гипометилирования DAC в различных клеточных линиях. После обработки DAC в течение 4 дней клетки собирали и выделяли ДНК. Метилирование LINE измеряли с помощью анализа бисульфитного пиросеквенирования. В каждой клеточной линии, кроме наиболее устойчивых клеток (нижний график), кривая зависимости от дозы имела U-образную форму. (B) Отсутствие корреляции IC 50 DAC с метилированием LINE на исходном уровне (R = 0,05, P = 0,97). (C) Корреляция между IC 50 DAC с дозами DAC, необходимыми для максимального гипометилирования LINE (R = 0,94, P <0,001).
Close modal
При поиске механизмов устойчивости к DAC мы сначала спросили, коррелирует ли она с метилированием ДНК. Глобальное метилирование с использованием повторяющегося элемента ДНК LINE в качестве маркера измеряли с помощью анализа на основе пиросеквенирования. Наблюдались большие различия в метилировании LINE на исходном уровне (от 12% в K562 до 79%). % в клеточной линии MOLT4), как сообщалось ранее 25 (таблица 1). Затем мы обрабатывали все клеточные линии DAC в дозах 0,03, 0,1, 0,3, 1, 3, 10 и 30 мкМ ежедневно в течение 4 дней и измеряли индукцию гипометилирования с помощью анализа LINE. В каждой клеточной линии ответ на дозу гипометилирования имел U-образную форму, как сообщалось ранее, 5 , предположительно потому, что высокие дозы DAC ингибируют пролиферацию, что необходимо для гипометилирования. Дозы DAC, вызывающие пиковое гипометилирование, варьировали от 0,03 мкМ в U9 до37 и TF-1, наиболее чувствительные клеточные линии; до 0,3 мкМ в чувствительных клеточных линиях HEL, Raji, ML-1, HL60, SW48 и Cama-1; до 1 и 3 мкМ в относительно устойчивых клеточных линиях Jurkat и MOLT4 соответственно; и до 30 мкМ в наиболее устойчивых клеточных линиях, RKO и PC3 (рис. 1А). IC 50 DAC тесно коррелировал с дозами, вызывающими пиковое гипометилирование LINE (R = 0,94, P < 0,001), но не с метилированием LINE на исходном уровне (R = 0,05, P = 0,97; Рисунок 1Б,С).
% в клеточной линии MOLT4), как сообщалось ранее 25 (таблица 1). Затем мы обрабатывали все клеточные линии DAC в дозах 0,03, 0,1, 0,3, 1, 3, 10 и 30 мкМ ежедневно в течение 4 дней и измеряли индукцию гипометилирования с помощью анализа LINE. В каждой клеточной линии ответ на дозу гипометилирования имел U-образную форму, как сообщалось ранее, 5 , предположительно потому, что высокие дозы DAC ингибируют пролиферацию, что необходимо для гипометилирования. Дозы DAC, вызывающие пиковое гипометилирование, варьировали от 0,03 мкМ в U9 до37 и TF-1, наиболее чувствительные клеточные линии; до 0,3 мкМ в чувствительных клеточных линиях HEL, Raji, ML-1, HL60, SW48 и Cama-1; до 1 и 3 мкМ в относительно устойчивых клеточных линиях Jurkat и MOLT4 соответственно; и до 30 мкМ в наиболее устойчивых клеточных линиях, RKO и PC3 (рис. 1А). IC 50 DAC тесно коррелировал с дозами, вызывающими пиковое гипометилирование LINE (R = 0,94, P < 0,001), но не с метилированием LINE на исходном уровне (R = 0,05, P = 0,97; Рисунок 1Б,С). Затем мы измерили экспрессию белков DNMT1, 3a и 3b с помощью вестерн-блоттинга. Белок DNMT1 зависит от клеточной репликации. Клетки в фазе быстрого роста экспрессировали больше белка DNMT1, чем в фазе медленного роста (рис. 2А). Например, в RKO уровень DNMT1 был высоким в 1-й день и постепенно снижался на 3-й и 5-й дни. Мы обнаружили, что экспрессия белков DNMT1, 3a и 3b в логарифмически растущих клетках не коррелировала с метилированием LINE на исходном уровне (рис. 2B). ) и не коррелировал с IC 50 ЦАП. Рисунок 2 (A) Экспрессия белка DNMT1 зависела от репликации клеток. Мы измерили кривую роста клеток RKO и экспрессию белка DNMT1 в дни 1, 3 и 5 с помощью вестерн-блоттинга. В качестве контроля использовали β-актин. (B) Экспрессия белков DNMT1, 3a и 3b не зависела от чувствительности к метилированию DAC и LINE в различных линиях раковых клеток. Мы собрали экспоненциально растущие раковые клетки, извлекли белок и провели вестерн-блоттинг DNMT1, 3a и 3b. В качестве контроля использовали β-актин.
Затем мы измерили экспрессию белков DNMT1, 3a и 3b с помощью вестерн-блоттинга. Белок DNMT1 зависит от клеточной репликации. Клетки в фазе быстрого роста экспрессировали больше белка DNMT1, чем в фазе медленного роста (рис. 2А). Например, в RKO уровень DNMT1 был высоким в 1-й день и постепенно снижался на 3-й и 5-й дни. Мы обнаружили, что экспрессия белков DNMT1, 3a и 3b в логарифмически растущих клетках не коррелировала с метилированием LINE на исходном уровне (рис. 2B). ) и не коррелировал с IC 50 ЦАП. Рисунок 2 (A) Экспрессия белка DNMT1 зависела от репликации клеток. Мы измерили кривую роста клеток RKO и экспрессию белка DNMT1 в дни 1, 3 и 5 с помощью вестерн-блоттинга. В качестве контроля использовали β-актин. (B) Экспрессия белков DNMT1, 3a и 3b не зависела от чувствительности к метилированию DAC и LINE в различных линиях раковых клеток. Мы собрали экспоненциально растущие раковые клетки, извлекли белок и провели вестерн-блоттинг DNMT1, 3a и 3b. В качестве контроля использовали β-актин. (C) Экспрессия белка dCK в нескольких клеточных линиях. Экспрессию белка dCK измеряли вестерн-блоттингом. (D) Корреляция экспрессии генов метаболизма различных нуклеозидов с IC 50 ЦАП. Экспрессию DCK , CDA , hENT1 и hENT2 измеряли с помощью ПЦР в реальном времени с использованием глицеральдегид-3-фосфатдегидрогеназы (GAPDH) в качестве контроля. Значения R и P отражают корреляционный анализ Спирмена IC 50 DAC с относительной экспрессией генов. Рисунок 2 (A) Экспрессия белка DNMT1 зависела от репликации клеток. Мы измерили кривую роста клеток RKO и экспрессию белка DNMT1 в дни 1, 3 и 5 с помощью вестерн-блоттинга. В качестве контроля использовали β-актин. (B) Экспрессия белков DNMT1, 3a и 3b не зависела от чувствительности к метилированию DAC и LINE в различных линиях раковых клеток. Мы собрали экспоненциально растущие раковые клетки, извлекли белок и провели вестерн-блоттинг DNMT1, 3a и 3b. В качестве контроля использовали β-актин.
(C) Экспрессия белка dCK в нескольких клеточных линиях. Экспрессию белка dCK измеряли вестерн-блоттингом. (D) Корреляция экспрессии генов метаболизма различных нуклеозидов с IC 50 ЦАП. Экспрессию DCK , CDA , hENT1 и hENT2 измеряли с помощью ПЦР в реальном времени с использованием глицеральдегид-3-фосфатдегидрогеназы (GAPDH) в качестве контроля. Значения R и P отражают корреляционный анализ Спирмена IC 50 DAC с относительной экспрессией генов. Рисунок 2 (A) Экспрессия белка DNMT1 зависела от репликации клеток. Мы измерили кривую роста клеток RKO и экспрессию белка DNMT1 в дни 1, 3 и 5 с помощью вестерн-блоттинга. В качестве контроля использовали β-актин. (B) Экспрессия белков DNMT1, 3a и 3b не зависела от чувствительности к метилированию DAC и LINE в различных линиях раковых клеток. Мы собрали экспоненциально растущие раковые клетки, извлекли белок и провели вестерн-блоттинг DNMT1, 3a и 3b. В качестве контроля использовали β-актин. (C) Экспрессия белка dCK в нескольких клеточных линиях. Экспрессию белка dCK измеряли вестерн-блоттингом. (D) Корреляция экспрессии генов метаболизма различных нуклеозидов с IC 50 ЦАП. Экспрессию DCK , CDA , hENT1 и hENT2 измеряли с помощью ПЦР в реальном времени с использованием глицеральдегид-3-фосфатдегидрогеназы (GAPDH) в качестве контроля. Значения R и P отражают корреляционный анализ Спирмена IC 50 DAC с относительной экспрессией генов.
(C) Экспрессия белка dCK в нескольких клеточных линиях. Экспрессию белка dCK измеряли вестерн-блоттингом. (D) Корреляция экспрессии генов метаболизма различных нуклеозидов с IC 50 ЦАП. Экспрессию DCK , CDA , hENT1 и hENT2 измеряли с помощью ПЦР в реальном времени с использованием глицеральдегид-3-фосфатдегидрогеназы (GAPDH) в качестве контроля. Значения R и P отражают корреляционный анализ Спирмена IC 50 DAC с относительной экспрессией генов.
Close modal
Из-за наблюдаемой корреляции между IC50 50 DAC и Ara-C, которые имеют общие внутриклеточные метаболические пути, мы исследовали экспрессию нескольких генов, участвующих в метаболизме нуклеозидов. Экспрессия белка dCK была очень низкой в устойчивых к DAC клеточных линиях DU145 и Jurkat (рис. 2C), что было связано с низкой экспрессией мРНК dCK с помощью ПЦР в реальном времени (рис. 2D). ИС 50 DAC обратно коррелировал с экспрессией экспрессии мРНК dCK (R = -0,63, P = 0,038), а также показал тенденцию к обратной корреляции с мРНК переносчика нуклеозидов hENT1 (R = -0,54, P = 0,068; рис. 2D). мРНК hENT1 была самой низкой в DAC-резистентной клеточной линии MOLT4. DAC-инактивирующий фермент CDA мРНК был самым высоким в PC3 и DU145, соответственно. Таким образом, 4 из 5 клеточных линий, наиболее устойчивых к DAC, имеют измеримые изменения в одном из этих генов, в то время как ни одна из 9наиболее чувствительные клеточные линии имели такие изменения. Напротив, IC 50 AZA не коррелировало с экспрессией DCK , CDA , hENT1 и hENT2 соответственно (данные не показаны). Интересно, что клеточная линия Raji была устойчива к AZA, но чувствительна к DAC и Ara-C. Промотор DCK представляет собой богатую CpG область, которая может быть подавлена гиперметилированием ДНК. Затем мы измерили уровень метилирования DCK во всех этих клеточных линиях с помощью анализа пиросеквенирования, но не смогли обнаружить какого-либо аберрантного гиперметилирования даже в DU145 и Jurkat с самой низкой экспрессией гена (данные не показаны).
ИС 50 DAC обратно коррелировал с экспрессией экспрессии мРНК dCK (R = -0,63, P = 0,038), а также показал тенденцию к обратной корреляции с мРНК переносчика нуклеозидов hENT1 (R = -0,54, P = 0,068; рис. 2D). мРНК hENT1 была самой низкой в DAC-резистентной клеточной линии MOLT4. DAC-инактивирующий фермент CDA мРНК был самым высоким в PC3 и DU145, соответственно. Таким образом, 4 из 5 клеточных линий, наиболее устойчивых к DAC, имеют измеримые изменения в одном из этих генов, в то время как ни одна из 9наиболее чувствительные клеточные линии имели такие изменения. Напротив, IC 50 AZA не коррелировало с экспрессией DCK , CDA , hENT1 и hENT2 соответственно (данные не показаны). Интересно, что клеточная линия Raji была устойчива к AZA, но чувствительна к DAC и Ara-C. Промотор DCK представляет собой богатую CpG область, которая может быть подавлена гиперметилированием ДНК. Затем мы измерили уровень метилирования DCK во всех этих клеточных линиях с помощью анализа пиросеквенирования, но не смогли обнаружить какого-либо аберрантного гиперметилирования даже в DU145 и Jurkat с самой низкой экспрессией гена (данные не показаны). Кроме того, альтернативный сплайсинг DCK не наблюдался в устойчивых клонах (данные не представлены). Вполне вероятно, что сниженная экспрессия мРНК DCK в некоторых клеточных линиях, таких как DU145 или Jurkat, может быть вызвана другими механизмами, такими как деацетилирование гистонов или микроРНК, нацеленные на DCK .
Кроме того, альтернативный сплайсинг DCK не наблюдался в устойчивых клонах (данные не представлены). Вполне вероятно, что сниженная экспрессия мРНК DCK в некоторых клеточных линиях, таких как DU145 или Jurkat, может быть вызвана другими механизмами, такими как деацетилирование гистонов или микроРНК, нацеленные на DCK .
Данные, описанные выше, относятся к естественной резистентности к DAC (и AZA). Чтобы изучить эту проблему в модели индуцированной устойчивости, мы обработали HL60 DAC, отобрали устойчивые колонии (HL60R) в среде с метилцеллюлозой и нашли частоту устойчивости 1,2 × 10 −4 . Мы повторно посеяли 11 устойчивых к DAC колоний и обнаружили, что устойчивость была стабильной и могла сохраняться более 2 лет даже при культивировании в отсутствие DAC. Чтобы исследовать механизмы индуцированной устойчивости к DAC в этих клонах, мы измерили индукцию гипометилирования DAC LINE-повторяющихся элементов. Гипометилирование, вызванное низкими дозами DAC (0,2 и 2 мкМ) в клетках HL60D, почти исчезло в DAC-резистентных клетках (рис. 3А). Чтобы изучить, влияет ли эта устойчивость на индукцию экспрессии генов с помощью DAC, мы измерили мРНК для RIL, гена-супрессора опухоли, который замалчивается в HL60 за счет гиперметилирования ДНК. 5 Реактивация RIL с помощью DAC также ингибировалась в HL60R (рис. 3B). Напротив, AZA ингибирует рост клеток (рис. 3C) и индуцирует деметилирование LINE (рис. 3D) в HL60D и HL60R аналогичным образом.
3А). Чтобы изучить, влияет ли эта устойчивость на индукцию экспрессии генов с помощью DAC, мы измерили мРНК для RIL, гена-супрессора опухоли, который замалчивается в HL60 за счет гиперметилирования ДНК. 5 Реактивация RIL с помощью DAC также ингибировалась в HL60R (рис. 3B). Напротив, AZA ингибирует рост клеток (рис. 3C) и индуцирует деметилирование LINE (рис. 3D) в HL60D и HL60R аналогичным образом.
Рисунок 3
Просмотреть в большом формате Загрузить PPT
Дефицит dCK в резистентных клетках HL60R . (A) Индукция гипометилирования DAC в HL60D и резистентном к DAC HL60R. Мы обработали клетки DAC (0,2-50 мкМ) и измерили метилирование LINE с помощью анализа бисульфитного пиросеквенирования. (B) Экспрессия гена RIL. (C) Ингибирование AZA роста клеток. Клетки обрабатывали AZA (0,25-25 мкМ) и измеряли жизнеспособность клеток по исключению трипанового синего. (D) Гипометилирование LINE после лечения AZA. (E) Производство Ara-CTP, измеренное анализом ВЭЖХ с использованием [ 3 H] Ara-C в качестве субстрата терялся в HL60R-резистентных клетках. (F) активность dCK, измеренная по фосфорилированию [ 3 H]Ara-C в клеточных экстрактах, также была потеряна в резистентных клетках. (G) экспрессию белка dCK измеряли вестерн-блоттингом. β-актин служил контролем. (H) Экспрессию мРНК DCK измеряли с помощью количественной ПЦР.
(F) активность dCK, измеренная по фосфорилированию [ 3 H]Ara-C в клеточных экстрактах, также была потеряна в резистентных клетках. (G) экспрессию белка dCK измеряли вестерн-блоттингом. β-актин служил контролем. (H) Экспрессию мРНК DCK измеряли с помощью количественной ПЦР.
Рисунок 3
Просмотреть в большом формате Загрузить PPT
Дефицит dCK в резистентных клетках HL60R . (A) Индукция гипометилирования DAC в HL60D и резистентном к DAC HL60R. Мы обработали клетки DAC (0,2-50 мкМ) и измерили метилирование LINE с помощью анализа бисульфитного пиросеквенирования. (B) Экспрессия гена RIL. (C) Ингибирование AZA роста клеток. Клетки обрабатывали AZA (0,25-25 мкМ) и измеряли жизнеспособность клеток по исключению трипанового синего. (D) Гипометилирование LINE после лечения AZA. (E) Производство Ara-CTP, измеренное анализом ВЭЖХ с использованием [ 3 H] Ara-C в качестве субстрата терялся в HL60R-резистентных клетках. (F) активность dCK, измеренная по фосфорилированию [ 3 H]Ara-C в клеточных экстрактах, также была потеряна в резистентных клетках. (G) экспрессию белка dCK измеряли вестерн-блоттингом. β-актин служил контролем. (H) Экспрессию мРНК DCK измеряли с помощью количественной ПЦР.
(G) экспрессию белка dCK измеряли вестерн-блоттингом. β-актин служил контролем. (H) Экспрессию мРНК DCK измеряли с помощью количественной ПЦР.
Close modal
В резистентных клетках DAC индуцировал наибольшее гипометилирование при высокой дозе 50 мкМ, предполагая, что резистентность может быть связана с уменьшением включения в ДНК. В соответствии с этим мы обнаружили, что эти устойчивые клетки также устойчивы к Ara-C (данные не показаны), и предположили, что устойчивость была обусловлена недостаточным включением NA в ДНК. Поэтому мы затем измерили концентрацию Ara-CTP с помощью анализа на основе ВЭЖХ с использованием [ 3 H]Ara-C в качестве субстрата и обнаружили, что продукция Ara-CTP в устойчивых клонах, полученных из DAC, была в 50 раз ниже, чем в клетках дикого типа (рис. 3E). Активность dCK была в 10 раз ниже, чем в клетках дикого типа с использованием [ 3 H]Ara-C в качестве субстрата (рис. 3F). При поиске причин этой потери активности мы обнаружили, что экспрессия белка dCK была заметно снижена в HL60R (рис. 3G), но экспрессия мРНК DCK в HL60R была аналогична таковой в клеточных линиях дикого типа (рис. 3H). Таким образом, индуцированная устойчивость к DAC в HL60 коррелировала с потерей dCK.
При поиске причин этой потери активности мы обнаружили, что экспрессия белка dCK была заметно снижена в HL60R (рис. 3G), но экспрессия мРНК DCK в HL60R была аналогична таковой в клеточных линиях дикого типа (рис. 3H). Таким образом, индуцированная устойчивость к DAC в HL60 коррелировала с потерей dCK.
В поисках механизма потери dCK в устойчивых к HL60 клетках мы сначала секвенировали DCK из кДНК и геномной ДНК родительских клеток HL60, которые использовали для создания устойчивых клонов. Мы обнаружили гетерозиготную точечную мутацию в кодоне 98 (от ACA до AGA), что приводит к изменению Thr/Arg. Эта последовательность в родительских клетках HL60, обозначенных как HL60D, отсутствовала в другой партии клеток HL60, полученных от ATCC, обозначенных как HL60 (фиг. 4А). Кроме того, клетки как HL60D, так и HL60 имели гетерозиготные синонимичные SNP в кодоне 100 (от GCC до GCT). Мы обнаружили, что HL60D имеет преимущество в росте по сравнению с HL60, предполагая, что эта точечная мутация произошла в субпопуляции клеток HL60 (рис. 4B). Важно отметить, что ни точечная мутация, ни соседний SNP не были обнаружены более чем в 100 аллелях других исследованных клеточных линий и нормальных клеток. Затем мы секвенировали кДНК dCK и геномную ДНК в устойчивых клетках HL60R (полученных из HL60D) в поисках дополнительных мутаций. Среди 11 устойчивых клонов 3 секвенированных клона HL60R имели экзон 3 LOH аллеля дикого типа, оставляя после себя исключительно аллель, содержащий точечную мутацию AGA в кодоне 9.8 и GCC в кодоне 100. Восемь клонов имели гетерозиготную делецию в экзоне 1 между нуклеотидами 154 и 197 мРНК DCK , которая отменяла сайт ATG (таблица 2 и фигура 4). Чтобы определить, приводит ли эта гетерозиготная делеция к двуаллельной мутации, мы выполнили клонирование ТА (Invitrogen) и секвенирование кДНК DCK из экзона 1-4. Мы обнаружили 2 мутантных аллеля DCK : аллель с точечной мутацией в кодоне 98 и еще один аллель с делецией 42 п.н. в экзоне 1. Чтобы определить, были ли мутации спонтанными или существовавшими ранее, мы разработали праймеры для ПЦР, охватывающие удаленную область DCK , экзон 1, и успешно амплифицируют эту область из родительских клеток HL60D (рис.
4B). Важно отметить, что ни точечная мутация, ни соседний SNP не были обнаружены более чем в 100 аллелях других исследованных клеточных линий и нормальных клеток. Затем мы секвенировали кДНК dCK и геномную ДНК в устойчивых клетках HL60R (полученных из HL60D) в поисках дополнительных мутаций. Среди 11 устойчивых клонов 3 секвенированных клона HL60R имели экзон 3 LOH аллеля дикого типа, оставляя после себя исключительно аллель, содержащий точечную мутацию AGA в кодоне 9.8 и GCC в кодоне 100. Восемь клонов имели гетерозиготную делецию в экзоне 1 между нуклеотидами 154 и 197 мРНК DCK , которая отменяла сайт ATG (таблица 2 и фигура 4). Чтобы определить, приводит ли эта гетерозиготная делеция к двуаллельной мутации, мы выполнили клонирование ТА (Invitrogen) и секвенирование кДНК DCK из экзона 1-4. Мы обнаружили 2 мутантных аллеля DCK : аллель с точечной мутацией в кодоне 98 и еще один аллель с делецией 42 п.н. в экзоне 1. Чтобы определить, были ли мутации спонтанными или существовавшими ранее, мы разработали праймеры для ПЦР, охватывающие удаленную область DCK , экзон 1, и успешно амплифицируют эту область из родительских клеток HL60D (рис. 4D), тем самым демонстрируя, что резистентность существовала ранее и была спонтанной. Рис. 4 (A) Два субклона клеточной линии HL60. HL60D развил гетерозиготную точечную мутацию 454C>G DCK . Эта мутация отсутствовала в другой партии HL60, полученной от ATCC. (B) Клеточная линия HL60D имела преимущество в росте по сравнению с клеточной линией HL60. Количество клеток подсчитывали с 1 по 7 день. (C) Гетерозиготная делеция в экзоне 1 из DCK в ячейках HL60R. (D) Делеция уже существовала в клетках HL60D и отсутствовала в клетках HL60. Мы разработали набор праймеров, которые охватывают удаленную область и амплифицируют эту область из родительских клеток HL60D. (E) Клетки HL60R, трансфицированные DCK , восстановили чувствительность к DAC. Мы трансфицировали DCK дикого типа в HL60R и отобрали стабильно трансфицированные клетки с помощью G418. Клетки, трансфицированные HL60R и DCK (HL60T), обрабатывали DAC в концентрациях 0,02, 0,2, 2 и 20 мкМ соответственно.
4D), тем самым демонстрируя, что резистентность существовала ранее и была спонтанной. Рис. 4 (A) Два субклона клеточной линии HL60. HL60D развил гетерозиготную точечную мутацию 454C>G DCK . Эта мутация отсутствовала в другой партии HL60, полученной от ATCC. (B) Клеточная линия HL60D имела преимущество в росте по сравнению с клеточной линией HL60. Количество клеток подсчитывали с 1 по 7 день. (C) Гетерозиготная делеция в экзоне 1 из DCK в ячейках HL60R. (D) Делеция уже существовала в клетках HL60D и отсутствовала в клетках HL60. Мы разработали набор праймеров, которые охватывают удаленную область и амплифицируют эту область из родительских клеток HL60D. (E) Клетки HL60R, трансфицированные DCK , восстановили чувствительность к DAC. Мы трансфицировали DCK дикого типа в HL60R и отобрали стабильно трансфицированные клетки с помощью G418. Клетки, трансфицированные HL60R и DCK (HL60T), обрабатывали DAC в концентрациях 0,02, 0,2, 2 и 20 мкМ соответственно. Подсчитывали жизнеспособность клеток. (F) Трансфекция 9кДНК 0014 DCK восстанавливала экспрессию белка dCK. Экспрессию белка dCK измеряли в клетках HL60R и HL60T с помощью вестерн-блоттинга. Рис. 4 (A) Два субклона клеточной линии HL60. HL60D развил гетерозиготную точечную мутацию 454C>G DCK . Эта мутация отсутствовала в другой партии HL60, полученной от ATCC. (B) Клеточная линия HL60D имела преимущество в росте по сравнению с клеточной линией HL60. Количество клеток подсчитывали с 1 по 7 день. (C) Гетерозиготная делеция в экзоне 1 из DCK в ячейках HL60R. (D) Делеция уже существовала в клетках HL60D и отсутствовала в клетках HL60. Мы разработали набор праймеров, которые охватывают удаленную область и амплифицируют эту область из родительских клеток HL60D. (E) Клетки HL60R, трансфицированные DCK , восстановили чувствительность к DAC. Мы трансфицировали DCK дикого типа в HL60R и отобрали стабильно трансфицированные клетки с помощью G418. Клетки, трансфицированные HL60R и DCK (HL60T), обрабатывали DAC в концентрациях 0,02, 0,2, 2 и 20 мкМ соответственно.
Подсчитывали жизнеспособность клеток. (F) Трансфекция 9кДНК 0014 DCK восстанавливала экспрессию белка dCK. Экспрессию белка dCK измеряли в клетках HL60R и HL60T с помощью вестерн-блоттинга. Рис. 4 (A) Два субклона клеточной линии HL60. HL60D развил гетерозиготную точечную мутацию 454C>G DCK . Эта мутация отсутствовала в другой партии HL60, полученной от ATCC. (B) Клеточная линия HL60D имела преимущество в росте по сравнению с клеточной линией HL60. Количество клеток подсчитывали с 1 по 7 день. (C) Гетерозиготная делеция в экзоне 1 из DCK в ячейках HL60R. (D) Делеция уже существовала в клетках HL60D и отсутствовала в клетках HL60. Мы разработали набор праймеров, которые охватывают удаленную область и амплифицируют эту область из родительских клеток HL60D. (E) Клетки HL60R, трансфицированные DCK , восстановили чувствительность к DAC. Мы трансфицировали DCK дикого типа в HL60R и отобрали стабильно трансфицированные клетки с помощью G418. Клетки, трансфицированные HL60R и DCK (HL60T), обрабатывали DAC в концентрациях 0,02, 0,2, 2 и 20 мкМ соответственно. Подсчитывали жизнеспособность клеток. (F) Трансфекция 9кДНК 0014 DCK восстанавливала экспрессию белка dCK. Экспрессию белка dCK измеряли в клетках HL60R и HL60T с помощью вестерн-блоттинга.
Подсчитывали жизнеспособность клеток. (F) Трансфекция 9кДНК 0014 DCK восстанавливала экспрессию белка dCK. Экспрессию белка dCK измеряли в клетках HL60R и HL60T с помощью вестерн-блоттинга.
Близкий модальный
Таблица 2
Резюме устойчивости к DAC в клетках HL60D
| Группа . | Отобранные устойчивые клоны . | Частота сопротивления ЦАП . | Мутация . | ||
|---|---|---|---|---|---|
| ЛОХ . | Удаление . | Сдвиг рамы . | |||
| HL6O | 0 | Undetected | 0 | 0 | 0 |
| HL6OD (pool) | 11 | 1..jpg) 2 × 10 −4 2 × 10 −4 | 3 | 8 | 0 |
| HL6OD (клон) | 9 | 1,2 × 10 −5 | 7 | 0 | 2 |
| Группа . | Отобранные устойчивые клоны . | Частота сопротивления ЦАП . | Мутация . | ||
|---|---|---|---|---|---|
| ЛОХ . | Удаление . | Сдвиг рамы . | |||
| HL6O | 0 | Undetected | 0 | 0 | 0 |
| HL6OD (pool) | 11 | 1.2 × 10 −4 | 3 | 8 |  “> 0 “> 0 |
| HL6OD (clone | 9 | 1,2 × 10 −5 | 7 | 0 | 2 |
. колонии HL60D. После того, как каждая колония увеличилась примерно до 10 6 , мы отобрали DAC-устойчивые клоны в среде с метилцеллюлозой и получили 9 устойчивых клонов. Средняя частота резистентности в этих клетках составила 1,2 × 10 –5 (табл. 2). Мы измерили аллельный статус DCK с помощью пиросеквенирования с использованием геномной ДНК и обнаружили, что из 9 одиночных устойчивых колоний 7 имели LOH DCK , что привело к единственному мутантному аллелю с Arg в кодоне 98 в экзоне 3. Два клона имели вставка одного основания, вызывающая сдвиг рамки (TA/TAA) в экзоне 6 из ДСК (табл. 2). Клонирование кДНК и секвенирование подтвердили, что в каждом случае мутация со сдвигом рамки считывания была в аллеле, не мутированном в экзоне 3. Таким образом, LOH DCK является наиболее распространенным механизмом, вызывающим индуцированную устойчивость к DAC в HL60. Чтобы подтвердить, что устойчивость вызвана потерей DCK , мы клонировали DCK дикого типа в векторы pEGFP-N1 и трансфицировали вектор в HL60R. Были отобраны стабильно устойчивые к G418 клетки, обозначенные как HL60T. Трансфекция DCK восстановил экспрессию белка dCK и чувствительность к DAC (рис. 4E, F), предполагая, что резистентность к DAC была обусловлена функциональной потерей dCK, которая была вызвана LOH локуса DCK .
Чтобы подтвердить, что устойчивость вызвана потерей DCK , мы клонировали DCK дикого типа в векторы pEGFP-N1 и трансфицировали вектор в HL60R. Были отобраны стабильно устойчивые к G418 клетки, обозначенные как HL60T. Трансфекция DCK восстановил экспрессию белка dCK и чувствительность к DAC (рис. 4E, F), предполагая, что резистентность к DAC была обусловлена функциональной потерей dCK, которая была вызвана LOH локуса DCK .
LOH может быть спонтанным или может быть индуцирован повреждением ДНК. Мы пришли к выводу, что LOH в клетках HL60R может быть связан с разрывами ДНК, вызванными DAC. Поэтому мы измерили индукцию фосфорилирования гистона h3AX после обработки DAC клеток HL60D. Вестерн-блоттинг показал, что обработка DAC в концентрации 0,2 и 2 мкМ индуцировала фосфорилирование h3AX (рис. 5А). Фосфорилирование, вызванное более высокой концентрацией, исчезало на 3-й день, возможно, из-за гибели клеток, тогда как более низкая концентрация поддерживала это фосфорилирование на 3-й день. LOH в аутосомном локусе является одним из результатов репарации двухцепочечных разрывов ДНК с помощью HRR. Чтобы выяснить, увеличивает ли лечение DAC скорость HRR, мы провели ретровирусную трансфекцию вектора PLNCX-GZ 9.0692 24 в ячейки HL60D. В этом анализе рекомбинация между тандемными повторами регенерирует функциональный маркер GFP-ZeoR, который можно легко оценить. Скорость HRR клеток HL60D составляла 1,4 × 10 -6 . Лечение 0,02, 0,2, 2 и 20 мкМ DAC повышало частоту ЧСС примерно в 1,9, 6,8, 9,5 и 4,1 раза соответственно (рис. 5B). Чтобы подтвердить, что устойчивые колонии были рекомбинантными, мы выделили высокомолекулярную ДНК из размноженных G418 плюс устойчивых к зеоцину колоний и провели ПЦР с праймерами, фланкирующими 2 кассеты GFP-зеоцин, которые могут идентифицировать ожидаемые события рекомбинации. Мы случайным образом выбрали 6 колоний после отбора зеоцином плюс G418 и отметили, что все колонии имели фрагмент размером 1,1 т.п.н., подтверждающий рекомбинацию.
LOH в аутосомном локусе является одним из результатов репарации двухцепочечных разрывов ДНК с помощью HRR. Чтобы выяснить, увеличивает ли лечение DAC скорость HRR, мы провели ретровирусную трансфекцию вектора PLNCX-GZ 9.0692 24 в ячейки HL60D. В этом анализе рекомбинация между тандемными повторами регенерирует функциональный маркер GFP-ZeoR, который можно легко оценить. Скорость HRR клеток HL60D составляла 1,4 × 10 -6 . Лечение 0,02, 0,2, 2 и 20 мкМ DAC повышало частоту ЧСС примерно в 1,9, 6,8, 9,5 и 4,1 раза соответственно (рис. 5B). Чтобы подтвердить, что устойчивые колонии были рекомбинантными, мы выделили высокомолекулярную ДНК из размноженных G418 плюс устойчивых к зеоцину колоний и провели ПЦР с праймерами, фланкирующими 2 кассеты GFP-зеоцин, которые могут идентифицировать ожидаемые события рекомбинации. Мы случайным образом выбрали 6 колоний после отбора зеоцином плюс G418 и отметили, что все колонии имели фрагмент размером 1,1 т.п.н., подтверждающий рекомбинацию. С другой стороны, колонии после селекции G418 имели только некомбинированный фрагмент размером 2,2 т.п.н. (фиг. 5C). 9Рис. 5 (A) Индукция фосфорилирования гистона h3AX обработкой DAC. Клетки HL60D обрабатывали DAC (0,2 и 2 мкМ) в течение 3 дней и фосфорилирование h3AX измеряли вестерн-блоттингом. В качестве контроля использовали β-актин. (B) Лечение DAC увеличило частоту ЧСС. Клетки со стабильной трансфекцией PLNCX-GZ обрабатывали DAC в течение 4 дней и выдерживали в среде без лекарств в течение 1 недели. Колонии, устойчивые к зеоцину, отбирали и выдерживали в среде с метилцеллюлозой в течение 2 недель. Показатели HRR рассчитывали, как описано в разделе «Измерение гомологичной рекомбинационной репарации». (C) Проверка внутрихромосомной рекомбинации в устойчивых к зеоцину колониях с помощью ПЦР с рекомбинационно-специфическими праймерами. ПЦР-амплифицированная ДНК из устойчивых к G418 колоний продуцировала фрагмент размером 2,2 т.п.н., диагностический для нерекомбинированных тандемных повторов.
С другой стороны, колонии после селекции G418 имели только некомбинированный фрагмент размером 2,2 т.п.н. (фиг. 5C). 9Рис. 5 (A) Индукция фосфорилирования гистона h3AX обработкой DAC. Клетки HL60D обрабатывали DAC (0,2 и 2 мкМ) в течение 3 дней и фосфорилирование h3AX измеряли вестерн-блоттингом. В качестве контроля использовали β-актин. (B) Лечение DAC увеличило частоту ЧСС. Клетки со стабильной трансфекцией PLNCX-GZ обрабатывали DAC в течение 4 дней и выдерживали в среде без лекарств в течение 1 недели. Колонии, устойчивые к зеоцину, отбирали и выдерживали в среде с метилцеллюлозой в течение 2 недель. Показатели HRR рассчитывали, как описано в разделе «Измерение гомологичной рекомбинационной репарации». (C) Проверка внутрихромосомной рекомбинации в устойчивых к зеоцину колониях с помощью ПЦР с рекомбинационно-специфическими праймерами. ПЦР-амплифицированная ДНК из устойчивых к G418 колоний продуцировала фрагмент размером 2,2 т.п.н., диагностический для нерекомбинированных тандемных повторов. ПЦР-амплифицированная ДНК из колоний, устойчивых к G418 и зеоцину, генерировала диагностический фрагмент размером 1,1 т.п.н. 9Рис. 5 (A) Индукция фосфорилирования гистона h3AX обработкой DAC. Клетки HL60D обрабатывали DAC (0,2 и 2 мкМ) в течение 3 дней и фосфорилирование h3AX измеряли вестерн-блоттингом. В качестве контроля использовали β-актин. (B) Лечение DAC увеличило частоту ЧСС. Клетки со стабильной трансфекцией PLNCX-GZ обрабатывали DAC в течение 4 дней и выдерживали в среде без лекарств в течение 1 недели. Колонии, устойчивые к зеоцину, отбирали и выдерживали в среде с метилцеллюлозой в течение 2 недель. Показатели HRR рассчитывали, как описано в разделе «Измерение гомологичной рекомбинационной репарации». (C) Проверка внутрихромосомной рекомбинации в устойчивых к зеоцину колониях с помощью ПЦР с рекомбинационно-специфическими праймерами. ПЦР-амплифицированная ДНК из устойчивых к G418 колоний продуцировала фрагмент размером 2,2 т.п.н., диагностический для нерекомбинированных тандемных повторов.
ПЦР-амплифицированная ДНК из колоний, устойчивых к G418 и зеоцину, генерировала диагностический фрагмент размером 1,1 т.п.н. 9Рис. 5 (A) Индукция фосфорилирования гистона h3AX обработкой DAC. Клетки HL60D обрабатывали DAC (0,2 и 2 мкМ) в течение 3 дней и фосфорилирование h3AX измеряли вестерн-блоттингом. В качестве контроля использовали β-актин. (B) Лечение DAC увеличило частоту ЧСС. Клетки со стабильной трансфекцией PLNCX-GZ обрабатывали DAC в течение 4 дней и выдерживали в среде без лекарств в течение 1 недели. Колонии, устойчивые к зеоцину, отбирали и выдерживали в среде с метилцеллюлозой в течение 2 недель. Показатели HRR рассчитывали, как описано в разделе «Измерение гомологичной рекомбинационной репарации». (C) Проверка внутрихромосомной рекомбинации в устойчивых к зеоцину колониях с помощью ПЦР с рекомбинационно-специфическими праймерами. ПЦР-амплифицированная ДНК из устойчивых к G418 колоний продуцировала фрагмент размером 2,2 т.п.н., диагностический для нерекомбинированных тандемных повторов. ПЦР-амплифицированная ДНК из колоний, устойчивых к G418 и зеоцину, генерировала диагностический фрагмент размером 1,1 т.п.н.
ПЦР-амплифицированная ДНК из колоний, устойчивых к G418 и зеоцину, генерировала диагностический фрагмент размером 1,1 т.п.н.
Close modal
Наконец, мы исследовали, была ли быстрая индукция устойчивости к DAC обычным явлением или это была специфическая черта клеток HL60D, гетерозиготных по мутации DCK . Обработка DAC и отбор метилцеллюлозы не дали стабильных резистентных клонов в клетках HL60 с двумя аллелями дикого типа DCK или в ML-1, клеточной линии с нормальной dCK.
В этом исследовании мы исследовали естественную резистентность к DAC in vitro на панели линий раковых клеток и обнаружили 1000-кратную разницу в IC 50 значения. Эти различия, вероятно, связаны с фармакологическими механизмами, приводящими к более низкому включению DAC в ДНК в резистентных клетках. Следовательно, для индукции максимального гипометилирования в резистентных клетках требовались более высокие дозы DAC. Кроме того, мы обнаружили, что как спонтанная генетическая нестабильность, так и генетическая нестабильность, индуцированная DAC, способствовали возникновению устойчивости к DAC в клетках HL60D, несущих миссенс-точечную мутацию в экзоне 3 DCK .
Большая часть устойчивости in vitro к NA возникает из-за отсутствия включения в ДНК. 26 В соответствии с этим мы обнаружили, что устойчивость к DAC коррелирует с несколькими фармакологическими механизмами аберрантного метаболизма нуклеозидов, что приводит к меньшему включению DAC в ДНК, но это не было напрямую доказано путем измерения включения DAC. Экспериментально очень трудно ответить на этот вопрос, потому что включение DAC находится на низком уровне. DAC имеет короткий период полураспада (минуты) и быстро дезаминируется CDA внутри клеток. Он также химически нестабилен при экстракции ДНК и анализе ВЭЖХ. Связь 5,6 в цитозине очень лабильна. 27,28 Тем не менее, в HL60 мы продемонстрировали, что резистентность к DAC вызывается потерей DCK , а чувствительность к DAC сильно коррелирует с чувствительностью к Ara-C, который имеет общий метаболизм с NA. Эксперименты с AraCTP являются косвенными, но они являются очень хорошим приближением активности dCK. Таким образом, данные в пользу инкорпорации как механизма устойчивости убедительны. Ясно, однако, что несколько механизмов могут быть активны в разных клетках.
Таким образом, данные в пользу инкорпорации как механизма устойчивости убедительны. Ясно, однако, что несколько механизмов могут быть активны в разных клетках.
DAC оказывает свое действие посредством индукции гипометилирования при низких дозах и цитотоксичности при высоких дозах. DAC индуцировал U-образную кривую гипометилирования в группе линий раковых клеток. Это может быть объяснено тем фактом, что низкие дозы DAC могут ковалентно улавливать ДНК-метилтрансферазы без остановки клеточного цикла, тогда как высокие дозы DAC ингибируют синтез ДНК и вызывают остановку клеточного цикла, что приводит к меньшей индукции гипометилирования. Однако клиническое использование агента в основном связано с низкими дозами. 4 Интересно, что устойчивость к DAC не была связана с уровнями DNMT и метилированием LINE, маркером глобального метилирования ДНК. Рак имеет геноспецифическое гиперметилирование и глобальное гипометилирование. 29 Наши данные свидетельствуют о том, что различия в уровнях метилирования повторяющихся элементов ДНК (отражающие глобальное метилирование) не просто объясняются уровнями DNMT и не влияют на чувствительность к DAC. Скорее индукция гипометилирования DAC и реактивация генов нарушаются в резистентных клетках из-за недостаточного включения DAC в ДНК. Потенциальный вопрос заключается в том, связаны ли нижестоящие пути, которые привели к реактивации генов, с устойчивостью к DAC, обеспечивающей адекватное включение лекарственного средства. Тот факт, что обработка DAC активирует тысячи генов, не все из которых подавляются гиперметилированием промотора, 30 затрудняет такое обучение. Однако в этой панели клеточных линий перекрестная устойчивость между DAC и Ara-C и отсутствие перекрестной устойчивости между DAC и AZA предполагает, что устойчивость к реактивации генов, возможно, менее важна, чем включение DAC. Тем не менее, устойчивость к DAC, вероятно, будет многофакторной в конкретной клеточной линии. Например, DU145 продемонстрировал низкие уровни DCK , hENT1 и высокие уровни CDA . Дозы, необходимые для максимального гипометилирования, являются лучшим маркером для определения чувствительности или устойчивости к DAC на основе наших результатов.
Скорее индукция гипометилирования DAC и реактивация генов нарушаются в резистентных клетках из-за недостаточного включения DAC в ДНК. Потенциальный вопрос заключается в том, связаны ли нижестоящие пути, которые привели к реактивации генов, с устойчивостью к DAC, обеспечивающей адекватное включение лекарственного средства. Тот факт, что обработка DAC активирует тысячи генов, не все из которых подавляются гиперметилированием промотора, 30 затрудняет такое обучение. Однако в этой панели клеточных линий перекрестная устойчивость между DAC и Ara-C и отсутствие перекрестной устойчивости между DAC и AZA предполагает, что устойчивость к реактивации генов, возможно, менее важна, чем включение DAC. Тем не менее, устойчивость к DAC, вероятно, будет многофакторной в конкретной клеточной линии. Например, DU145 продемонстрировал низкие уровни DCK , hENT1 и высокие уровни CDA . Дозы, необходимые для максимального гипометилирования, являются лучшим маркером для определения чувствительности или устойчивости к DAC на основе наших результатов.
В наших исследованиях мы смогли объяснить устойчивость двумя потенциальными механизмами: (1) существующей генетической нестабильностью и отбором устойчивых клонов и (2) генетической нестабильностью, вызванной DAC. Большинство популяций злокачественных клеток характеризуются «генетической нестабильностью», которая, как можно показать, непосредственно участвует в формировании фенотипической лекарственной устойчивости. Используя анализ флуктуаций, исследователи продемонстрировали случайное и спонтанное происхождение фенотипов устойчивости к некоторым противоопухолевым средствам, таким как ингибитор топоизомеразы II, антагонисты фолиевой кислоты и антибиотики. 10-16 В этом исследовании мы амплифицировали мутации, ранее существовавшие в редких резистентных клетках из пула родительских клеток, с помощью ПЦР, предоставив самое прямое доказательство спонтанного происхождения резистентности к DAC и другим NA. С другой стороны, NA влияют на структурную целостность ДНК, приводя к остановке вилок репликации и обрыву цепи. Датчики повреждения ДНК ATM, ATR и DNA-PK распознают эти события и сигнализируют о репарации ДНК. 31 Мы наблюдали, что обработка DAC индуцировала фосфорилирование гистона h3AX, маркер двухцепочечных разрывов ДНК, и повышала скорость HRR, что могло привести к LOH DCK и устойчивость к DAC. Некоторые цитостатические препараты, в том числе афидиколин, Ara-C, гидроксимочевина и метотрексат, индуцируют HRR посредством ингибирования синтеза ДНК. 32
Датчики повреждения ДНК ATM, ATR и DNA-PK распознают эти события и сигнализируют о репарации ДНК. 31 Мы наблюдали, что обработка DAC индуцировала фосфорилирование гистона h3AX, маркер двухцепочечных разрывов ДНК, и повышала скорость HRR, что могло привести к LOH DCK и устойчивость к DAC. Некоторые цитостатические препараты, в том числе афидиколин, Ara-C, гидроксимочевина и метотрексат, индуцируют HRR посредством ингибирования синтеза ДНК. 32
Чувствительность к DAC в наших исследованиях была тесно связана с чувствительностью к Ara-C, но не к AZA, за исключением клеточной линии PC3, которая имела высокие уровни CDA , способные дезаминировать все эти препараты в неактивные формы. DAC и Ara-C нуждаются в одном и том же ферменте dCK для начального фосфорилирования. Вместо этого AZA использует UCK для начального фосфорилирования. Следовательно, резистентность к DAC, связанная с потерей dCK, не приводит к резистентности к AZA, и это отсутствие перекрестной резистентности можно использовать в терапевтических целях. Действительно, недавнее исследование показало, что некоторые пациенты могут реагировать на DAC после проявления клинической резистентности к AZA. 33
Действительно, недавнее исследование показало, что некоторые пациенты могут реагировать на DAC после проявления клинической резистентности к AZA. 33
Необходимо изучить актуальность наших выводов относительно резистентности in vivo. Низкие уровни экспрессии гена DCK или низкая активность dCK коррелировали со слабым ответом на Ara-C и кладрибин (2-хлордеоксиаденозин) при остром лимфобластном лейкозе (ALL) и лимфопролиферативных заболеваниях у детей, 34,35 и на гемцитабин ( аналог цитозина) при распространенном раке поджелудочной железы. 36 Однако не все исследования были последовательными в этом отношении. 37,38 Наши объединенные данные показывают, что лучшим подходом к изучению устойчивости in vivo к DAC было бы измерение включения DAC в ДНК, эксперимент, который в настоящее время технически неосуществим из-за низких доз DAC, используемых в клинике. Альтернативным подходом может быть измерение всех параметров метаболизма DAC (например, dCK, hENT, CDA) у пролеченных пациентов и их корреляция с чувствительностью к препарату.
В заключение, это исследование предоставляет важные модели in vitro для понимания механизмов и происхождения устойчивости к DAC, что может привести к стратегиям преодоления устойчивости к DAC in vivo. Отсутствие перекрестной резистентности между DAC и AZA в некоторых клетках также может иметь важные терапевтические последствия.
Затраты на публикацию этой статьи были частично покрыты оплатой постраничных расходов. Поэтому и исключительно для того, чтобы указать на этот факт, эта статья настоящим помечена как «реклама» в соответствии с разделом 1734 18 USC.
Мы благодарим доктора Уильяма Планкетта и Билли Новак за общие советы и помощь в измерении Ara-CTP и dCK. д-р Лу Сюнбин и Лоуренс А. Донехауэр (Медицинский колледж Бейлора, Хьюстон, Техас) за предоставление векторов PLNCX-GZ, pVSV-G и pHit60 для изучения гомологичной рекомбинации, а также д-р Гергели Кеслер из Университета Земмельвайса (Будапешт, Венгрия). ) для получения антитела dCK.
Эта работа была поддержана грантами Национального института здравоохранения CA100632 и CA108631, а также стипендией Розали Б. Хайт для выпускников Центра медицинских наук Техасского университета в Хьюстоне.
Хайт для выпускников Центра медицинских наук Техасского университета в Хьюстоне.
Национальные институты здравоохранения
Вклад: T.Q. проведенное исследование; Дж.Дж. и Ж.-П.Дж.И. проектное исследование; и Дж. Си и Дж. Шу проанализировали данные.
Раскрытие информации о конфликте интересов: Авторы заявляют об отсутствии конкурирующих финансовых интересов.
Для переписки: Жан-Пьер Дж. Исса, отделение лейкемии, Онкологический центр доктора медицины Андерсона Техасского университета, Хьюстон, Техас; электронная почта: [email protected].
1
Соларс
ВЭ
.
Эпигенетическая модификация как активирующий механизм лейкемической трансформации.
,
Front Biosci
,
2005
, том.
10
(pg.
1635
–
1646
)
2
Villa
R
,
De Santis
F
,
Gutierrez
A
,
Minucci
S
,
Pelicci
PG
,
Di Croce
L
.
Эпигенетическое замалчивание генов при остром промиелоцитарном лейкозе.
,
Biochem Pharmacol
,
2004
, том.
68
(стр.
1247
–
1254
)
3
Mizuno
S
,
Chijiwa
T
,
Okamura
T
, et al.
Экспрессия ДНК-метилтрансфераз DNMT1, 3A и 3B при нормальном кроветворении, остром и хроническом миелогенном лейкозе.
,
Кровь
,
2001
, том.
97
(стр.
1172
–
1179
)
4
ISSA
JP
.
Метилирование ДНК как терапевтическая мишень при раке.
,
Clin Cancer Res
,
2007
, том.
13
(стр.
1634
–
1637
)
5
90s 5 900ef 0 , 90ef0638EM
,
Елинек
J
, и др.
Влияние комбинации цитарабина и децитабина на лейкемические клеточные линии человека.
,
Clin Cancer Res
,
2007
, том.
13
(pg.
4225
–
4232
)
6
Lyko
F
,
Brown
Р
.
Ингибиторы ДНК-метилтрансферазы и разработка эпигенетических методов лечения рака.
,
J Natl Cancer Inst
,
2005
, vol.
97
(pg.
1498
–
1506
)
7
Kroep
JR
,
Loves
WJ
,
Ван дер Вилт
CL
, и др.
Уровни дезоксицитидинкиназы до лечения позволяют прогнозировать чувствительность к гемцитабину in vivo.
,
Мол Рак Тер
,
2002
, том.
1
(pg.
371
–
376
)
8
Flasshove
M
,
Strumberg
D
,
Ayscue
L
, и др.
Структурный анализ гена дезоксицитидинкиназы у больных острым миелоидным лейкозом и резистентностью к цитозинарабинозиду.
,
Лейкемия
,
1994
, том.
8
(стр.
780
–
785
)
Stegmann
AP
0638 ,
Honders
MW
,
Hagemeijer
A
,
Hoebee
B
,
Willemze
R
,
Landegent
JE
.
Индуцированная in vitro резистентность к аналогам дезоксицитидина цитарабину (AraC) и 5-аза-2′-дезоксицитидину (DAC) в крысиной модели острого миелоидного лейкоза опосредована мутациями в гене дезоксицитидинкиназы (DCK).
,
Энн Хематол
,
1995
, том.
71
(pg.
41
–
47
)
10
Wang
G
,
Wilson
TJ
,
Jiang
Q
,
Тейлор
DE
.
Спонтанные мутации, придающие Helicobacter pylori устойчивость к антибиотикам.
,
Противомикробные агенты Chemother
,
2001
, vol.
45
(pg.
727
–
733
)
11
Vogler
AJ
,
Busch
JD
,
Percy-Fine
S
,
Типтон-Хантон
C
, 9.
Молекулярный анализ устойчивости к рифампину Bacillus anthracis и Bacillus cereus.
,
Противомикробные агенты Chemother
,
2002
, том.
46
(стр.
511
–
513
)
12
9 600005
9 0 8 0 0
JP
,
Chen
G
,
Duran
GE
,
Kuhl
JS
,
Sikic
BI
.
Частота мутаций и механизмы устойчивости к этопозиду, определенные на основе анализа флуктуаций.
,
J Natl Cancer Inst
,
1994
, vol.
86
(pg.
1152
–
1158
)
13
Poche
H
,
Varshaver
NB
,
Geissler
Е
.
Устойчивость к циклогексимиду в клетках китайских хомячков. I. Спонтанный мутагенез.
,
Мутат Рез
,
1975
, том.
27
(стр.
399
–
406
)
14
GOLDIE
JH
,
9898988888888888888888888888888888888888888888888888888888888888888888888888888888898988 9000 9000 9000.
Генетическая нестабильность в развитии лекарственной устойчивости.
,
Семин Онкол
,
1985
, том.
12
(стр.
222
–
230
)
15
Моннат
RJ
.
Молекулярный анализ спонтанных мутаций гипоксантинфосфорибозилтрансферазы в тиогуанин-резистентных клетках лейкемии человека HL-60.
,
Рак Рез
,
1989
, том.
49
(стр.
81
–
87
)
16
Siminovitch
L
.0005.
О характере наследственной изменчивости культивируемых соматических клеток.
,
Сотовый
,
1976
, том.
7
(pg.
1
–
11
)
17
Chen
RZ
,
Pettersson
U
,
Beard
С
,
Джексон-Грусби
L
,
Jaenisch
R
.
Гипометилирование ДНК приводит к повышенной частоте мутаций.
,
Природа
,
1998
, том.
395
(стр.
89
–
93
)
18
Lengauer
C
,
Kinzl0638
кВт
,
Фогельштейн
B
.
Генетическая нестабильность при раке человека.
,
Природа
,
1998
, том.
396
(pg.
643
–
649
)
19
Raderschall
E
,
Stout
K
,
Freier
S
,
Suckow
V
,
Schweiger
S
,
Haaf
T
.
Повышенные уровни рекомбинационного белка Rad51 в опухолевых клетках.
,
Рак Рез
,
2002
, том.
62
(стр.
219
–
225
)
20
,
Hernandez
R
,
Zhang
xy
, ET.
Деметилирование ДНК и перицентромерные реаранжировки хромосомы 1.
379
(стр.
33
–
41
)
21
9.Количественный анализ зависимости «доза-эффект»: комбинированное действие нескольких лекарств или ингибиторов ферментов.
,
Adv Enzyme Regul
,
1984
, vol.
22
(стр.
27
–
55
)
22
58 Родригес
80005
CO
,
Планкетт
W
,
Paff
,
MTal 5. et al.
Метод высокоэффективной жидкостной хроматографии для определения и количественного определения арабинозилгуанинтрифосфата и флударабинтрифосфата в клетках человека.
,
J Chromatogr B Biomed Sci Appl
,
2000
, vol.
745
(стр.
421
–
430
)
23
Saunders
PP
,
LAI
MM
.
Активность нуклеозидкиназы клеток яичника китайского хомячка.
,
Биохим Биофиз Акта
,
1983
, том.
761
(стр.
135
–
141
)
24
,
Лосано
G
,
Донехауэр
LA
.
Активность р53 дикого типа и мутанта в подавлении гомологичной рекомбинации, измеренная с помощью ретровирусной векторной системы.
,
Мутат Рез
,
2003
, том.
522
(стр.
69
–
83
)
25
Estecio
MR
,
Gharibyan
V
,
Shen
L
, et.
Гипометилирование LINE-1 при раке сильно варьирует и обратно коррелирует с микросателлитной нестабильностью.
,
PLoS ONE
,
2007
, том.
2
стр.
e399
26
Galmarini
см
,
Mackey
JR
,
Dumontet
C
.
Аналоги нуклеозидов: механизмы лекарственной устойчивости и стратегии устранения.
,
Лейкемия
,
2001
, том.
15
(стр.
875
–
890
)
27
Момпарлер
RL
.
Фармакология 5-аза-2′-дезоксицитидина (децитабина).
,
Семин Гематол
,
2005
, том.
42
(pg.
S9
–
S16
)
28
Yoo
CB
,
Jeong
S
,
Egger
G
и др.
Доставка 5-аза-2′-дезоксицитидина в клетки с использованием олигодезоксинуклеотидов.
,
Рак Рез
,
2007
, том.
67
(стр.
6400
–
6408
)
29
Робертсон
KD
.
Метилирование ДНК: прошлое, настоящее и будущее.
,
Канцерогенез
,
2000
, том.
21
(pg.
461
–
467
)
30
Schmelz
K
,
Sattler
N
,
Wagner
M
,
Любберт
M
,
Доркен
Б
,
Тамм
I
.
Индукция экспрессии генов 5-аза-2′-дезоксицитидином при остром миелоидном лейкозе (ОМЛ) и миелодиспластическом синдроме (МДС), но не в эпителиальных клетках, посредством ДНК-метилирования-зависимых и независимых механизмов.
,
Лейкемия
,
2005
, том.
19
(стр.
103
–
111
)
31
SAMPATH
D
,
RAO
VA
,
.
Механизмы индукции апоптоза аналогами нуклеозидов.
,
Онкоген
,
2003
, том.
22
(стр.
9063
–
9074
)
32
Arnaudeau
C
,
Tenorio
ME
,
Jenssen
D
,
Helleday
T
.
Ингибирование синтеза ДНК является мощным механизмом, с помощью которого цитостатические препараты вызывают гомологичную рекомбинацию в клетках млекопитающих.
,
Мутат Рез
,
2000
, том.
461
(pg.
221
–
228
)
33
Borthakur
G
,
Ahdab
SE
,
Ravandi
F
и др.
Активность децитабина у пациентов с миелодиспластическим синдромом, ранее получавших азацитидин.
,
Лейк-лимфома
,
2008
, том.
49
(pg.
690
–
695
)
34
Kakihara
T
,
Fukuda
T
,
Tanaka
A
и др.
Экспрессия гена дезоксицитидинкиназы (dCK) в лейкозных клетках у детей: снижение экспрессии гена dCK при рецидиве лейкемии.
,
Лейк-лимфома
,
1998
, том.
31
(pg.
405
–
409
)
35
Stammler
G
,
Zintl
F
,
Sauerbrey
А
,
Том
М
.
Экспрессия мРНК дезоксицитидинкиназы при остром лимфобластном лейкозе у детей.
,
Противораковые препараты
,
1997
, том.
8
(pg.
517
–
521
)
36
Sebastiani
V
,
Ricci
F
,
Rubio-Viqueira
B
и др.
Иммуногистохимическая и генетическая оценка дезоксицитидинкиназы при раке поджелудочной железы: связь с молекулярными механизмами резистентности к гемцитабину и выживания.
,
Clin Cancer Res
,
2006
, том.
12
(pg.
2492
–
2497
)
37
Veuger
MJ
,
Honders
MW
,
Willemze
R
,
Баржа
RM
.
Экспрессия и активность дезоксицитидинкиназы у пациентов с резистентным и чувствительным острым миелоидным лейкозом.
,
Eur J Haematol
,
2002
, том.
69
(pg.
171
–
178
)
38
Leiby
JM
,
Snider
KM
,
Kraut
EH
,
Мец
EN
,
Мальспейс
L
,
Гревер
MR
.
Исследование фазы II 9-бета-D-арабинофуранозил-2-фтораденин 5′-монофосфата при неходжкинской лимфоме: проспективное сравнение ответа с активностью дезоксицитидинкиназы.
,
Рак Рез
,
1987
, том.
47
(стр.
2719
–
2722
)
Войдите через свое учреждение
ESS_5600Wilshire_SiteMap_Consolidated
%PDF-1.5 % 1 0 объект >/OCGs[10 0 R 11 0 R 12 0 R 13 0 R 14 0 R 15 0 R 175 0 R 176 0 R 177 0 R 178 0 R 179 0 R 180 0 R]>>/Страницы 3 0 R/Тип /Каталог>> эндообъект 2 0 объект >поток приложение/pdf
 did:14c9e9b7-642b-4e5c-a355-0e49da898617uuid:5D208BFDB11914A8590D31508C8proof:pdfuuid:0ec1fd96-9078-f441-9a4d-c5a1127b9407xmp.did:ff02a671-851f-4dd5-ab1b-b5b63c8cb6f5uuid: Adobe Illustrator 24.0 (Macintosh)/
did:14c9e9b7-642b-4e5c-a355-0e49da898617uuid:5D208BFDB11914A8590D31508C8proof:pdfuuid:0ec1fd96-9078-f441-9a4d-c5a1127b9407xmp.did:ff02a671-851f-4dd5-ab1b-b5b63c8cb6f5uuid: Adobe Illustrator 24.0 (Macintosh)/ 0000000.0000000.0000000.000000
0000000.0000000.0000000.000000NJDEP Департамент охраны окружающей среды Нью-Джерси
Сформированный штормом и приливами, государственный парк Айленд-Бич представляет собой узкий барьерный остров, протянувшийся на 10 миль между беспокойным Атлантическим океаном и историческим заливом Барнегат. Айленд-Бич – один из последних значительных остатков экосистемы барьерного острова в Нью-Джерси, которая когда-то существовала на большей части побережья, а также является одним из немногих оставшихся неосвоенных барьерных пляжей на побережье Северной Атлантики. Более 3000 акров и 10 миль прибрежных дюн остаются почти нетронутыми с тех пор, как Генри Хадсон впервые описал побережье Нью-Джерси с корабля «Полумесяц» в 1609 году..
Айленд-Бич – один из последних значительных остатков экосистемы барьерного острова в Нью-Джерси, которая когда-то существовала на большей части побережья, а также является одним из немногих оставшихся неосвоенных барьерных пляжей на побережье Северной Атлантики. Более 3000 акров и 10 миль прибрежных дюн остаются почти нетронутыми с тех пор, как Генри Хадсон впервые описал побережье Нью-Джерси с корабля «Полумесяц» в 1609 году..
Мили песчаных дюн и белых песчаных пляжей предлагают среду обитания для морских растений и разнообразной дикой природы, которая почти такая же, как и тысячи лет назад. Айленд-Бич содержит выдающиеся образцы растительных сообществ, таких как первичные дюны, заросли, пресноводные водно-болотные угодья, морские леса и приливные болота. Наш парк является домом для крупнейшей в штате колонии скоп, и они приветствуют самых разнообразных птиц, включая сапсанов, болотных, куликов, водоплавающих и перелетных певчих птиц. Айленд-Бич известен на национальном уровне как уникальный ресурс с идентифицированным более 400 растений, включая самые большие просторы пляжной погоды в Нью-Джерси.
Поддерживайте чистоту в парке
В рамках программы Carry-In/Carry-Out вы можете помочь нам сохранить парки чистыми и красивыми, вынося мусор, который вы приносите. Пожалуйста, возьмите с собой биоразлагаемый пакет, чтобы забрать мусор домой. Спасибо за сотрудничество и не забывайте перерабатывать.
Доступ для лиц с ограниченными возможностями
Удобства государственного парка Айленд-Бич частично доступны для людей с ограниченными возможностями. Рыбацкая дорожка — это дощатый настил, который обеспечивает доступ как к океану, так и к заливу в зоне парковки 7. Пляжные инвалидные коляски для зоны плавания в океане и пляжные инвалидные коляски, специально приспособленные для серфинга, доступны в павильонах во время купального сезона и в офис парка в оставшуюся часть года. Свяжитесь с офисом парка для получения дополнительной информации о доступе для людей с ограниченными возможностями. Пользователи текстовых телефонов (TT), позвоните в службу ретрансляции Нью-Джерси по телефону (800) 852-789. 9.
9.
Информация о парках
Указ губернатора о возобновлении работы парков
- Открытие парков в субботу, 2 мая
- Только пассивное использование
- Требуется социальное дистанцирование
- Покрытие лица настоятельно рекомендуется
- Парковка снижена на 50%
Посетите https://njparksandforests.org/sps_covid.html для получения самой последней информации о работе парка — некоторые удобства в настоящее время недоступны.
МЫ ВСЕ В ЭТОМ ВМЕСТЕ
Чтобы обезопасить свою семью и всю нашу семью в Нью-Джерси, оставайтесь дома, соблюдайте правила гигиены и соблюдайте все рекомендации штата и CDC в отношении COVID-19.
ПОДРОБНЕЕ
Чтобы узнать больше о COVID-19 или найти ресурсы, перейдите на https://www.COVID19.NJ.GOV, отправьте сообщение NJCOVID на номер 898-211 или позвоните по номеру 211.
Для получения обновленной информации о парках посетите сайт www.facebook.com/newjerseystateparks
– или –
Посетите веб-сайт Отдела парков и лесного хозяйства: https://njparksandforests. org
org
Часы работы парка
- Часы работы парка: 8:00–20:00
- Время рыбалки: с 5:00 до 20:00.
- Туалеты в зоне плавания 1: с 9:00 до 18:00.
- Все остальные туалеты закрыты.
- Офис закрыт.
Новости парка
Пляжная камера на OBA-1 предоставлена The Friends of Island Beach State Park
Консультации по навесу палатки
Посетите «оригинальный» берег Джерси, нетронутый и нетронутый – Государственный парк Айленд-Бич, показанный в шоу WNEP Home and Backyard
Щелкните здесь для просмотра видео
Карта парка
Карта государственного парка Айленд-Бич
Маршруты
Карта маршрутов по пляжу острова (файл PDF, 1 МБ)
Маршруты по пляжу острова — общее руководство
Брошюра о водной тропе морской заповедной зоны островов Седж
Система маршрутов Discovery Trails в парке штата Айленд-Бич обеспечивает самостоятельную экскурсию по девяти растительным сообществам барьерного острова. Придорожные экспонаты, расположенные вдоль каждой тропы, интерпретируют природные и культурные истории парка. Всего здесь 8 трасс, каждая меньше километра. Чтобы получить копию брошюры и карты Discovery Trails, обратитесь в офис парка.
Придорожные экспонаты, расположенные вдоль каждой тропы, интерпретируют природные и культурные истории парка. Всего здесь 8 трасс, каждая меньше километра. Чтобы получить копию брошюры и карты Discovery Trails, обратитесь в офис парка.
Верховая езда разрешена в государственном парке Айленд-Бич с 1 октября по 30 апреля. В южной и центральной частях парка есть шесть миль океанского пляжа, доступного для верховой езды, и в это время для конных трейлеров выделена стоянка. период. Для бронирования необходимо обратиться в офис парка по телефону (732) 79.3-0506.
Зона отдыха
Десять миль пляжа с чистым белым песком привлекают купающихся, загорающих, рыболовов и серферов. Современные бани, пандусы для доступа к пляжу, миля пляжа со спасателями, исторические здания, тропы, программы перевода, велосипедные дорожки и удобства для людей с ограниченными возможностями в совокупности делают Island Beach уникальным местом для изучения.
Залив Барнегат
Залив острова является богатым питательными веществами местом кормления птиц, рыб и других диких животных. В летние месяцы туры на каноэ и байдарках с гидом-натуралистом проходят через острова Седж, где они могут наблюдать за разнообразной дикой природой, в том числе за гнездящимися скопами, соколами и болотными птицами. Жалюзи для наблюдения за птицами дают посетителям возможность наблюдать за дикой природой в этой очень важной среде обитания.
В летние месяцы туры на каноэ и байдарках с гидом-натуралистом проходят через острова Седж, где они могут наблюдать за разнообразной дикой природой, в том числе за гнездящимися скопами, соколами и болотными птицами. Жалюзи для наблюдения за птицами дают посетителям возможность наблюдать за дикой природой в этой очень важной среде обитания.
Островной пляж Северная природная зона (659 акров) и Южная природная зона (1237 акров)
Вместе эти две природные территории составляют один из немногих оставшихся неосвоенных барьерных пляжей на северо-востоке США. Катящиеся песчаные дюны, соляная растительность, пышные зеленые солончаки и девять растительных сообществ, включая морской лес национального значения, характеризуют Айленд-Бич. Находящийся под угрозой исчезновения зуек-зубчик является случайным обитателем открытых пляжей и дюн. Доступ к частям Северной природной зоны ограничен.
Заявка на получение разрешения на специальное использование
Обратите внимание: Разрешение на специальное использование подлежит рассмотрению и утверждению парком.
Пакет заявок на получение разрешения на специальное использование
Разрешение на специальное использование используется для размещения определенной деятельности или мероприятия, проводимого в течение короткого периода времени. Существует два типа разрешений на специальное использование: некоммерческое и коммерческое, а сборы зависят от резидентства штата Нью-Джерси и нерезидентов. Заполненную заявку необходимо подать в парковую/лесную зону, где проводится мероприятие или мероприятие, не менее 90 дней до мероприятия. Если специальное использование или событие чрезвычайно большое или сложное, рекомендуется предварительное уведомление не менее чем за год.
Домашние животные
С выходных, посвященных Дню поминовения, и до первого октября домашние животные не допускаются на пляжи для купания в океане (зоны, охраняемые спасателями). Домашние животные допускаются к плаванию к югу от океана в течение этого периода времени, но их необходимо держать на поводке (максимальная длина — 6 футов), и вы должны убирать за своим питомцем. Домашние животные не допускаются на тропу для слепых птиц Спиззл-Крик в любое время.
Домашние животные не допускаются на тропу для слепых птиц Спиззл-Крик в любое время.
Рыбалка
Государственный парк Айленд-Бич хорошо известен рыбакам в морской воде Нью-Джерси благодаря отличной ловле полосатого окуня и луфаря прибоем. Другие виды включают летнюю камбалу и слабую рыбу. В соответствующий сезон особенно выгодна ловля таутог у северного причала залива Барнегат. Информацию о наживке и рыбалке можно получить у местных продавцов снастей перед входом в парк. Морская рыбалка регулируется правилами Управления рыбных ресурсов и дикой природы штата Нью-Джерси и Службы государственных парков. Информацию о правилах рыболовства в морской воде можно найти в дайджесте рыбы и дикой природы, который можно приобрести у местных дилеров снастей.
Чтобы ограничить распространение COVID-19, Mobile Sportfishing Vehicle разрешает использовать полноприводные автомобили на рыболовных пляжах только по почте. Скачать заявление на годовое разрешение на мобильную спортивную рыбалку
| Резидент Нью-Джерси $75,00 Нерезидент $90,00 | Разрешение на 3 дня |
| Резидент Нью-Джерси $195,00 Нерезидент $225,00 | Годовое разрешение |
| Плата за замену $25,00 | Годовое разрешение |
Выданная наклейка должна рассматриваться как наличные деньги. Он не будет заменен в случае потери или кражи.
Он не будет заменен в случае потери или кражи.
SPS продолжит предлагать сменную наклейку за 25 долларов США только при предъявлении оплаченного чека и старой наклейки, независимо от ее состояния.
Разрешение MSFV:
- Выдается только зарегистрированному владельцу транспортного средства — имя в лицензии и регистрации должно совпадать.
- Арендованные автомобили должны предоставить документы об аренде.
- Транспортные средства, принадлежащие компании, должны иметь документы, подтверждающие право собственности. Принимается визитная карточка, если на ней указано «владелец» или «президент».
- Все документы должны иметь текущую действительную дату.
ОБЯЗАТЕЛЬНОЕ ОБОРУДОВАНИЕ ДЛЯ МОБИЛЬНОГО СПОРТИВНО-РЫБОЛОВНОГО АВТОМОБИЛЯ (MSFV)
Разрешения выдаются владельцам полноприводных транспортных средств только для рыболовных целей. Перевозка перечисленного основного оборудования требуется всякий раз, когда MSFV находится на пляже.
- Рыболовное снаряжение, наживка и снасти на каждого человека старше 12 лет
- Шинный манометр
- Запасное колесо
- Рабочий домкрат и доска/опора для домкрата в песке (минимальный размер фанерного квадрата 3/4 дюйма x 12 x 12 дюймов)
- Буксирная цепь или трос
- Лопата
- Фонарик
- Огнетушитель
- Автомобильная аптечка
- Мешок для мусора/мусора
- Минимум 1/4 бака топлива.
Текущая диаграмма приливов настоятельно рекомендуется. Также рекомендуются соединительные кабели для конкретного типа аккумуляторной батареи автомобиля.
Плавание
Закрыт на сезон.
Купание разрешено только тогда, когда дежурят спасатели с 10:00 до 18:00.
Приблизительно одна миля обозначенного пляжа для купания в океане расположена в центральной части парка. В период с середины июня до Дня труда на этом пляже для купания работают спасатели, обученные методам спасения жизни в океане и методам оказания первой помощи. Пожалуйста, ознакомьтесь с нашим расписанием плавания перед посещением. Рядом находятся парковки и банные павильоны с раздевалками, медпунктами и буфетами.
Пожалуйста, ознакомьтесь с нашим расписанием плавания перед посещением. Рядом находятся парковки и банные павильоны с раздевалками, медпунктами и буфетами.
Индивидуальные плавсредства не разрешается использовать в зонах купания в океане.
В этом летнем сезоне рядом с автомагистралью A-7, к северу от точки входа на Рыбацкую дорожку, откроется новая зона для плавания под охраной спасателей. Плата за вход в размере 3 долларов США (бесплатно для детей в возрасте до 3 лет) будет взиматься с тех, кто входит в парк на Рыбацкой дорожке, прилегающей к Тайс-Шол, пока дежурят спасатели.
назад
Пикник
Хотя в парке нет столов и грилей, посетители могут устроить пикник на берегу океана. Грили можно использовать на пляже к югу от места для купания. Костры должны быть не менее чем в 50 футах к востоку от дюн, и участники пикника должны потушить костры, прежде чем покинуть это место. Государственный парк Айленд-Бич требует, чтобы посетители «выносили/выносили» свой мусор.
Езда на велосипеде и верховая езда
Обозначенная велосипедная дорожка проходит более чем в 8 милях от главной дороги. Шесть миль океанского пляжа в южной и центральной частях парка доступны для верховой езды с 1 октября по 30 апреля. В этот период для конных прицепов предназначена стоянка. В парке или рядом с ним нет пунктов проката лошадей. Пожалуйста, свяжитесь с администрацией парка для получения дополнительной информации.
Парусный спорт и серфинг
Части океанского пляжа в крайнем южном конце обозначенной зоны для купания были отведены для парусного спорта и серфинга. Для получения дополнительной информации, пожалуйста, свяжитесь с администрацией парка.
Дайвинг
Подводное плавание с аквалангом и подводная рыбалка разрешены вдоль 2,5 миль океанского пляжа к северу от залива Барнегат. Аквалангисты должны зарегистрироваться в офисе парка каждый год непосредственно перед своим первым погружением. Требуется подтверждение действующего сертификата дайвинга. Подводная и подводная рыбалка регулируется правилами Отдела охраны рыбных ресурсов и дикой природы штата Нью-Джерси. Дайвинг в заливе Барнегат разрешен и регулируется специальными правилами Комиссии по гребле Нью-Джерси.
Подводная и подводная рыбалка регулируется правилами Отдела охраны рыбных ресурсов и дикой природы штата Нью-Джерси. Дайвинг в заливе Барнегат разрешен и регулируется специальными правилами Комиссии по гребле Нью-Джерси.
Программы интерпретации
Естественные образования песчаных дюн и относительно нетронутая растительность в государственном парке Айленд-Бич создают уникальный опыт изучения экосистемы барьерного острова на открытом воздухе. Школьные группы, желающие посетить парк для самостоятельных экскурсий, должны сделать предварительный заказ. Для получения дополнительной информации, пожалуйста, позвоните в офис парка по телефону (732) 793-0506.
Посмотреть и зарегистрироваться на программы
Морская заповедная зона острова Седж
Морская заповедная зона острова Седж содержит около 1600 акров высокопродуктивных приливных болот, ручьев, прудов и открытой воды. Цель Заповедной зоны состоит в том, чтобы защищать и интерпретировать ресурсы дикой природы, уменьшая конфликты между пользователями и сохраняя традиционное использование территории. Переводчики парка организуют туры на каноэ и байдарках. Для получения дополнительной информации о заповедной зоне, пожалуйста, свяжитесь с администрацией парка.
Переводчики парка организуют туры на каноэ и байдарках. Для получения дополнительной информации о заповедной зоне, пожалуйста, свяжитесь с администрацией парка.
- Административный приказ о морской заповедной зоне острова Седж № 2014-09 (pdf)
- Карта морской заповедной зоны острова Седж
Интерпретационный центр Эмили ДеКэмп
Гербарий Эмили ДеКэмп представляет собой коллекцию из примерно 400 растений, которые были классифицированы и сохранены, что дает публике возможность узнать о девяти растительных сообществах в государственном парке Айленд-Бич. Одной из главных достопримечательностей этого гербария является то, что посетители и ученые могут обрабатывать и исследовать отдельные растения. Растения помещены в защитную подставку, которая также дает историческую и фактическую информацию о растении. Гербарий хранится в Интерпретационном центре Форкед-Ривер.
Другие особенности
Две станции береговой охраны с 1901 по 1938 год; Крупнейшая колония скоп в Нью-Джерси.