Технические характеристики
|
СРАВНЕНИЕ |
КОМПЛЕКТ ПОСТАВКИ |
|
ТРЕБОВАНИЯ |
СХЕМА |
СРАВНЕНИЕ С ДРУГИМИ НАСОСАМИ-ДОЗАТОРАМИ
АСДР “Комплексон®-6”:
– работает в автоматическом режиме;
– оборудование занимает мало места;
– реагентов расходуется в сотни раз меньше, чем соли;
– полностью отсутствуют собственные сточные воды;
– не требуется постоянный лабораторный контроль, т.к. персонал котельной контролирует работу установки по имеющимся на ней приборам;
– реагенты имеют санитарно-эпидемиологические заключения и могут применяться для ГВС, открытых систем теплоснабжения и при подготовке питьевой воды.
В любой водооборотной системе не бывает идеально устойчивого давления, будь то трубопровод подпитки, система отопления, охлаждения и т. д. Поэтому насос-дозатор (НД), используемый для дозирования реагентов в данных системах должен иметь стабильную дозировку во всём диапазоне рабочих давлений.
д. Поэтому насос-дозатор (НД), используемый для дозирования реагентов в данных системах должен иметь стабильную дозировку во всём диапазоне рабочих давлений.
К сожалению, многие производители НД умалчивают данные о зависимости дозы НД от давления воды в точке ввода реагента. А это очень важный параметр, без которого нельзя быть уверенным в точности дозирования реагентов и, соответственно, в надёжности работы водооборотной системы в целом.
На графике хорошо видно – насколько сильно изменяется дозировка даже у именитого зарубежного производителя насосов-дозаторов, и это при применении электроники для выравнивания дозировки!
Большинство импортных НД спроектированы для работы при напряжении 230V ±10%, а то и 230V ±5%, такие требования по питанию не могут быть достигнуты, учитывая, что нормальное напряжение в Российской сети 220V ±10%, причём и оно не всегда соответствует этому диапазону. АСДР “Комплексон®-6″ специально разработана для эксплуатации в широком диапазоне напряжений питания и работает без изменения заявленных характеристик при питании 220V ±15%. Постоянное совершенствование химводоподготовки ООО «Дикма» позволило достичь эксклюзивных показателей стабильности дозирования. Поэтому АСДР «Комплексон®‑6» не имеет аналогов и остаётся вне конкуренции.
Постоянное совершенствование химводоподготовки ООО «Дикма» позволило достичь эксклюзивных показателей стабильности дозирования. Поэтому АСДР «Комплексон®‑6» не имеет аналогов и остаётся вне конкуренции.
КОМПЛЕКТ ПОСТАВКИ И ТЕХНИЧЕСКИЕ ХАРАКТЕРИСТИКИ
Электронный блок управления, насос-дозатор, расходная емкость, водосчетчик с импульсным выходом, устройство ввода реагента, монтажный комплект армированного шланга и провода, разовая заправка реагентом Эктоскейл-450-1 (цинковый комплекс НТФ 20%-ный раствор), комплект техдокументации.
| Расход подпиточной воды | Габаритный размер Ш*Г*В |
Комплект поставки | |||
| наминальный (усредненный) |
максимальный | ДУ водосчетчика |
Объём расходной емкости, л |
Разовая заправка реагентом, кг |
|
| 0,5 м3/час*3 |
2 м3/час | 460*190*340 | 15*2 |
5 | 6 |
| 1,5 м3/час*3 |
4 м3/час | 330*290*800 | 20*2 |
25 | 36 |
| 5 м3/час*3 |
10 м3/час | 470*510*950 | 32*2 |
60 | 36 |
| 10 м3/час*3 |
20 м3/час | 480*520*1140 | 50 | 100 | 72 |
| 20 м 3/час*3 |
40 м3/час | 570*610*1380 | 65 | 200 | 144 |
| 40 м3/час*3 |
80 м3/час | 940*940*1220 | 80 | 500 | 288 |
| 80 м3/час*3 |
160 м3/час | 1350*1350*1280 | 150 | 1000 | 360 |
| Более 80 м3/час*3 |
Для больших объёмов подпитки |
||||
*1 ДУ водосчётчик может изменятся по желанию заказчика
*2 В комплектацию входит фильтр грязевик
| Напряжение питания | 220 V ±15% |
| Средняя потребляемая мощность | 30 Вт |
| Максимальное давление воды в точке ввода реагента | стандартное исполнение – 0,8 МПа (8 кгс/см2) |
| по заказу – 1,2 МПа (12 кгс/см2) | |
| Предельный перепад давления на узле измерения и впрыска | 0,1 МПа (1,0 кгс/см2) |
| Основная приведенная погрешность дозирования при номинальных параметрах | менее 0,5% |
| Основная приведенная погрешность дозирования при колебаниях давления в системе ±0,2 МПа (2 кгс/см2) | ±2% |
| Насос – дозатор | мембранный с электромагнитным приводом |
| Клапан | спаренные, самоочищающиеся |
| Срок эксплуатации | не менее 10 лет |
ООО “Дикма” гарантирует
соответствие АСДР “Комплексон®”
требованиям паспорта
и ТУ 3631-001-31599189-96,
ТУ 3632-001-72582580-2016.
Декларация о соответствии
ЕАЭС № RU Д-RU.РА01.В.58478/21
Требования к размещению и монтажу АСДР “Комплексон®-6”
Водосчётчик входит в комплект поставки и снабжен электрическим адаптером для передачи информации о величине расхода и устройством ввода реагента (реагент вводится из блока управления и дозирования (БУД) непосредственно в водосчётчик или фильтр-грязевик и смешивается в нем с подпиточной водой).
1. Водосчётчик монтируется на трубопроводе подпитки и должен иметь байпас для возможности ремонта или замены.
2. Сама АСДР “Комплексон®-6″ размещается в любом удобном для обслуживания месте (но лучше поближе к водосчётчику) и соединяется с водосчётчиком двужильным проводом, а с узлом впрыска реагента – гибким армированным шлангом (Øнар=11мм). По проводу адаптер водосчётчика подает на БУД сигнал о величине расхода подпиточной воды, а по армированному шлангу в систему впрыскивается расчётное количество реагента и смешивается с подпиточной водой.
3. К АСДР “Комплексон®-6″ никакие трубопроводы не подходят. Рядом с ней должна быть розетка 220 вольт и доступ для заправки реагента в расходную емкость. Т.к. сама установка занимает мало места, ее обычно можно разместить недалеко от водосчётчика.
Если места не хватает, можно разместить в другом помещении, желательно не далее 20-30 метров от водосчётчика.
4. Контроль за работой АСДР “Комплексон®-6″ ведется по показаниям водосчётчика, поэтому его лучше разместить так, чтобы можно было удобно снимать показания (невысоко от пола и достаточное освещение). Таким образом, монтаж АСДР “Компдексон®-6″ сводится к монтажу водосчётчика.
5. Монтаж БУДа представляет собой соединение его двужильным проводом с водосчётчиком, гибким армированным шлангом с узлом впрыска реагента, заправке реагентом и прокачке насоса-дозатора от воздуха.
6. АСДР “Комплексон®-6″ поставляются настроенными по дозированию и после монтажа для включения в работу необходимо подать напряжение 220 вольт и установка начинает работать. Если через водосчётчик начинает протекать подпиточная вода, сигнал о величине расхода подается на БУД, обрабатывается и в соответствии с настройками в подпиточную воду через узел впрыска реагента вводится заданное количество реагента. Чем больше расход воды, тем больше подача реагента, нет расхода воды – нет подачи реагента. Таким образом, при любом расходе воды выдерживается заданная концентрация реагента в подпиточной воде. В системе управления предусмотрены специальные блокировки, которые не позволяют передозировать реагент при любых неисправностях.
Если через водосчётчик начинает протекать подпиточная вода, сигнал о величине расхода подается на БУД, обрабатывается и в соответствии с настройками в подпиточную воду через узел впрыска реагента вводится заданное количество реагента. Чем больше расход воды, тем больше подача реагента, нет расхода воды – нет подачи реагента. Таким образом, при любом расходе воды выдерживается заданная концентрация реагента в подпиточной воде. В системе управления предусмотрены специальные блокировки, которые не позволяют передозировать реагент при любых неисправностях.
Поэтому АСДР “Комплексон®-6″ имеет сертификат соответствия и её можно (в отличии от просто насосов-дозаторов) применять для ГВС, где передозировка реагента недопустима.
Продукция
Защита от накипи
Химическая деаэрация
Корректировка рН
Нестандартные решения
Реагенты
В НАЛИЧИИ
Расход подпиточной воды при О2=10 мг/дм3 – 1,5 м3/час
Расход подпиточной воды при О2=5 мг/дм3 – 3 м3/час
В комплекте с водосчетчиком ДУ-20
Габаритные размеры 470*510*950 (Ш*Г*В)
ПОДРОБНЕЕ . ..
..
В НАЛИЧИИ
Расход подпиточной воды при О2=10 мг/дм3 – 4 м3/час
Расход подпиточной воды при О2=5 мг/дм3 – 8 м3/час
В комплекте с водосчетчиком ДУ-32
Габаритные размеры 570*610*1380 (Ш*Г*В)
ПОДРОБНЕЕ …
В НАЛИЧИИ
Расход подпиточной воды при О2=10 мг/дм3 – 8 м3/час
Расход подпиточной воды при О2=5 мг/дм3 – 16 м3/час
В комплекте с водосчетчиком ДУ-40
Габаритные размеры 940*940*1220 (Ш*Г*В)
ПОДРОБНЕЕ …
В НАЛИЧИИ
Расход подпиточной воды при О2=10 мг/дм3 – 16 м3/час
Расход подпиточной воды при О2=5 мг/дм3 – 32 м3/час
В комплекте с водосчетчиком ДУ-50
Габаритные размеры 1350*1350*1280 (Ш*Г*В)
ПОДРОБНЕЕ . ..
..
В НАЛИЧИИ
(Цинковый комплекс НТФ 20%-ный водный раствор)
Канистры по 22 кг
ПОДРОБНЕЕ …
В НАЛИЧИИ
Подпитка усреднённая – 0,5 м3/час
Подпитка максимальная – 2 м3/час
В комплекте с водосчетчиком ДУ-15 и фильтром-“грязевиком”
Габаритные размеры – 460*190*340 (Ш*Г*В)
Возможен настенный монтаж
ПОДРОБНЕЕ …
В НАЛИЧИИ
Производительность насоса-дозатора – 2 л/час
Ориентировочный расход подпиточной воды – 2 м3/час
В комплекте с водосчетчиком ДУ-20 и фильтром-“грязевиком”
Габаритные размеры 470*510*950 (Ш*Г*В)
ПОДРОБНЕЕ …
В НАЛИЧИИ
Подпитка усреднённая – 1,5 м3/час
Подпитка максимальная – 4 м3/час
В комплекте с водосчетчиком ДУ-20 и фильтром-“грязевиком”
Габаритные размеры – 470*510*950 (Ш*Г*В)
ПОДРОБНЕЕ .
В НАЛИЧИИ
(Цинковый комплекс НТФ 20%-ный водный раствор)
Канистры по 6 кг
ПОДРОБНЕЕ …
В НАЛИЧИИ
Производительность насоса-дозатора – 2,5 л/час
Ориентировочный расход подпиточной воды – 5 м3/час
В комплекте с водосчетчиком ДУ-32 и фильтром-“грязевиком”
Габаритные размеры 570*610*1380 (Ш*Г*В)
ПОДРОБНЕЕ …
В НАЛИЧИИ
Производительность насоса-дозатора – 5 л/час
Ориентировочный расход подпиточной воды – 10 м3/час
В комплекте с водосчетчиком ДУ-50
Габаритные размеры 940*940*1220 (Ш*Г*В)
ПОДРОБНЕЕ …
В НАЛИЧИИ
(Цинковый комплекс НТФ)
Мешки по 25 кг
ПОДРОБНЕЕ …
В НАЛИЧИИ
Подпитка усреднённая – 5 м3/час
Подпитка максимальная – 10 м3/час
В комплекте с водосчетчиком ДУ-32
Габаритные размеры – 470*510*950 (Ш*Г*В)
Возможен настенный монтаж
ПОДРОБНЕЕ . ..
..
В НАЛИЧИИ
Производительность насоса-дозатора – 10 л/час
Ориентировочный расход подпиточной воды – 20 м3/час
В комплекте с водосчетчиком ДУ-65
Габаритные размеры 1350*1350*1280 (Ш*Г*В)
ПОДРОБНЕЕ …
В НАЛИЧИИ
(Цинковый комплекс НТФ 20%-ный водный раствор)
Канистры по 36 кг
ПОДРОБНЕЕ …
В НАЛИЧИИ
Подпитка усреднённая – 10 м3/час
Подпитка максимальная – 20 м3/час
В комплекте с водосчетчиком ДУ-50
Габаритные размеры – 480*520*1140 (Ш*Г*В)
ПОДРОБНЕЕ …
В НАЛИЧИИ
(Цинковый комплекс НТФ 20%-ный водный раствор)
ПОДРОБНЕЕ …
В НАЛИЧИИ
(Цинковый комплекс НТФ)
Мешки по 20 кг
ПОДРОБНЕЕ …
В НАЛИЧИИ
Подпитка усреднённая – 20 м3/час
Подпитка максимальная – 40 м3/час
В комплекте с водосчетчиком ДУ-65
Габаритные размеры – 570*610*1380 (Ш*Г*В)
ПОДРОБНЕЕ . ..
..
В НАЛИЧИИ
Мешки по 25 кг
ПОДРОБНЕЕ …
В НАЛИЧИИ
Подпитка усреднённая – 40 м3/час
Подпитка максимальная – 80 м3/час
В комплекте с водосчетчиком ДУ-80
Габаритные размеры 940*940*1220 (Ш*Г*В)
ПОДРОБНЕЕ …
В НАЛИЧИИ
Мешки по 25 кг
ПОДРОБНЕЕ …
В НАЛИЧИИ
Подпитка усреднённая – 80 м3/час
Подпитка максимальная –
В комплекте с водосчетчиком ДУ-150
Габаритные размеры 1350*1350*1280 (Ш*Г*В)
ПОДРОБНЕЕ …
Metro Parks открывает новый тепличный комплекс на полигоне Такома
Современное сооружение, которое, как ожидается, обеспечит долгосрочную экономию воды, энергии и обслуживания В.В. Ботаническая консерватория Сеймура в водовороте красного, зеленого, белого и розового цветов.
Эффектная экспозиция под названием «Роскошная» является праздничной традицией в элегантной оранжерее в викторианском стиле на восточной стороне Райт-парка.
Разреженная атмосфера Консерватории идеально подходит для пуансеттии тропического происхождения. Но эти растения были выращены в другом месте. Практически все горшечные растения, представленные в рамках сезонного календаря выставок консерватории, выращиваются в теплицах, обычно закрытых для публики. Группа стареющих теплиц на восточной окраине парка Пойнт-Дефаенс служила этой цели более полувека.
Это начинает меняться. Старые теплицы будут демонтированы в рамках реконструкции набережной парка Пойнт-Дефаенс и 60-миллионного Суперфонда по очистке загрязненной почвы, оставленной бывшим плавильным заводом ASARCO в Растоне.
По соглашению с городскими властями Такомы на краю старой городской свалки ведется строительство меньшего по размеру, но более эффективного тепличного комплекса. Его центральным элементом является ультрасовременная производственная теплица площадью 6000 квадратных футов с автоматизированной системой климат-контроля, которая следит за погодой и соответствующим образом регулирует отопление, освещение и вентиляцию. По оценкам, здание сэкономит деньги Metro Parks на воду и энергию, а также на долгосрочное обслуживание.
По оценкам, здание сэкономит деньги Metro Parks на воду и энергию, а также на долгосрочное обслуживание.
В октябре садоводы Metro Parks и другие специалисты перенесли первые растения из старых теплиц в новые условия. Среди них около 1000 пуансеттий, многие из которых сейчас выставлены в консерватории, и, возможно, такое же количество небольших папоротников различных разновидностей.
Новое здание со стальным каркасом и окнами из теплоизолированного поликарбоната, чрезвычайно прочного пластика. Он заменяет 60-летние деревянные теплицы, пропускающие свет через стеклянные панели. Это была настоящая головная боль при обслуживании, сказал специалист по садоводству Скотт Бассетт. Например, когда требовалась покраска, рабочим приходилось снимать и заменять каждое стекло.
Рядом с новым зданием бригады Metro Parks перенесли стальной каркас со старого участка, чтобы возвести менее сложные теплицы с теплым каркасом и затененную зону. Завершение всего проекта запланировано на ближайшие несколько месяцев. Деньги на объекты идут от $198 миллионов вопрос облигации одобренный избирателями парков Metro в 2014.
Деньги на объекты идут от $198 миллионов вопрос облигации одобренный избирателями парков Metro в 2014.
Двигать парники от пункта неповиновения было предметом обсуждения на около 20 лет. «Я никогда не думала, что это произойдет, пока мы не приступили к делу», — сказала Тайра Шенаурлт, руководитель садоводческих ресурсов Metro Parks. Она курирует и консерваторию, и оранжереи.
Одной из особенностей новой производственной теплицы, позволяющей сэкономить деньги, являются ее параллельные водоводы, которые отделяют удобренную воду от свежей питьевой воды. В старом тепличном комплексе рабочие должны были очищать удобренную воду из единственного водопровода, прежде чем его можно было использовать для орошения только водой. По словам администратора проекта Тома Даргана, эта модернизация может снизить потребление воды на 10–25 процентов.
Шенаурлт также рассчитывает на значительную общую экономию энергии. «Мы потеряли так много тепла в тех других теплицах», — сказала она.
Когда все будет готово, новый тепличный комплекс удовлетворит все садоводческие потребности Metro Parks. Помимо выращивания растений для консерватории, он будет выращивать однолетние растения, которые заполняют клумбы в парке Пойнт-Дефаенс, вдоль Растон-Уэй, а также в парках Вапато, Титлоу, Норпойнт и Дэш-Пойнт.
«Это действительно красивое место в центре», — сказал Бассет о новом тепличном кампусе. Это изменение потенциально может снизить стоимость транспортировки растений на выставочные площадки и обратно. В конце концов, часть нового кампуса можно было бы обозначить как карантинную зону для больных растений из консерватории или других мест.
На весну ориентировочно запланирован открытый день открытых дверей для нового тепличного комплекса; конкретная дата еще не назначена.
# # #
Контакты:
- Tyra Shenaurlt, W.W. Ботаническая консерватория Сеймура, (253) 591-5330; [электронная почта защищена]
Размещено в: Мест
Фосфопротеомика раскрывает сеть субстрата AMPK в ответ на повреждение ДНК и ацетилирование гистонов
1. Акланд М., Миттал П., Локман Н.А., Клинглер-Хоффманн М., Олер М.К., Хоффманн П. Масс-спектрометрический анализ сфероидов многоклеточных опухолей. Протеомика Clin Appl. 2018;12:e1700124. [PubMed] [Google Scholar]
Акланд М., Миттал П., Локман Н.А., Клинглер-Хоффманн М., Олер М.К., Хоффманн П. Масс-спектрометрический анализ сфероидов многоклеточных опухолей. Протеомика Clin Appl. 2018;12:e1700124. [PubMed] [Google Scholar]
2. Михайлова М.М., Шоу Р.Дж. Сигнальный путь AMPK координирует рост клеток, аутофагию и метаболизм. Nat Cell Biol. 2011;13:1016–1023. [Бесплатная статья PMC] [PubMed] [Google Scholar]
3. Shaw R.J., Kosmatka M., Bardeesy N., Hurley R.L., Witters L.A., DePinho R.A., et al. Киназа-супрессор опухолей LKB1 напрямую активирует AMP-активированную киназу и регулирует апоптоз в ответ на энергетический стресс. Proc Natl Acad Sci U S A. 2004; 101:3329–3335. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Форетц М., Гигас Б., Виоллет Б. От рака к лечению диабета: супрессор опухоли LKB1 как новая фармакологическая мишень. мед. наук. 2006; 22: 348–350. [PubMed] [Google Scholar]
5. Shaw R.J., Lamia K.A., Vasquez D., Koo S.H., Bardeesy N., Depinho R.A., et al. Киназа LKB1 опосредует гомеостаз глюкозы в печени и терапевтические эффекты метформина. Наука. 2005; 310:1642–1646. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Киназа LKB1 опосредует гомеостаз глюкозы в печени и терапевтические эффекты метформина. Наука. 2005; 310:1642–1646. [Бесплатная статья PMC] [PubMed] [Google Scholar]
6. Zhou G., Myers R., Li Y., Chen Y., Shen X., Fenyk-Melody J., et al. Роль АМФ-активируемой протеинкиназы в механизме действия метформина. Джей Клин Инвест. 2001; 108:1167–1174. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7. Сторожук Ю., Хопманс С.Н., Санли Т., Бэррон С., Циани Э., Катц Дж.К. и др. Метформин ингибирует рост и усиливает лучевую реакцию немелкоклеточного рака легкого (НМРЛ) посредством АТМ и АМРК. Бр Дж Рак. 2013;108:2021–2032. [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Sun Y., Tao C., Huang X., He H., Shi H., Zhang Q., et al. Метформин индуцирует апоптоз клеток гепатоцеллюлярной карциномы человека HepG2 путем активации пути AMPK/p53/miR-23a/FOXA1. Onco нацеливается на Ther. 2016;9:2845–2853. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Zou J., Hong L. , Luo C., Li Z., Zhu Y., Huang T., et al. Метформин ингибирует рост клеток эстрогензависимого рака эндометрия, активируя сигнальный путь AMPK-FOXO1. Онкологические науки. 2016; 107:1806–1817. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Luo C., Li Z., Zhu Y., Huang T., et al. Метформин ингибирует рост клеток эстрогензависимого рака эндометрия, активируя сигнальный путь AMPK-FOXO1. Онкологические науки. 2016; 107:1806–1817. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Чжуан Ю., Мискиминьш В.К. Остановка клеточного цикла в обработанных метформином клетках рака молочной железы включает активацию AMPK, подавление циклина D1 и требует p27Kip1 или p21Cip1. Дж Мол Сигнал. 2008;3:18. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Терапевтический потенциал метаболического модулятора метформина в отношении раковых стволовых клеток остеосаркомы. Рак Chemother Pharmacol. 2018;81:49–63. [PubMed] [Google Scholar]
12. Song C.W., Lee H., Dings R.P., Williams B., Powers J., Santos T.D., et al. Метформин убивает и делает радиосенсибилизированными раковые клетки и предпочтительно убивает раковые стволовые клетки. Научный доклад 2012; 2:362. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Soo J.S., Ng CH, Tan S.H., Malik R.A., Teh YC, Tan B.S., et al. Метформин синергизирует комбинированную терапию 5-фторурацилом, эпирубицином и циклофосфамидом (ФЭК), нарушая внутриклеточную выработку АТФ и репарацию ДНК в стволовых клетках рака молочной железы. Апоптоз. 2015;20:1373–1387. [PubMed] [Академия Google]
Soo J.S., Ng CH, Tan S.H., Malik R.A., Teh YC, Tan B.S., et al. Метформин синергизирует комбинированную терапию 5-фторурацилом, эпирубицином и циклофосфамидом (ФЭК), нарушая внутриклеточную выработку АТФ и репарацию ДНК в стволовых клетках рака молочной железы. Апоптоз. 2015;20:1373–1387. [PubMed] [Академия Google]
14. Sui X., Xu Y., Yang J., Fang Y., Lou H., Han W., et al. Использование только метформина не связано с выживаемостью клеток колоректального рака, но активатор AMPK AICAR сенсибилизирует противораковый эффект 5-фторурацила посредством активации AMPK. ПЛОС Один. 2014;9:e97781. [Бесплатная статья PMC] [PubMed] [Google Scholar]
15. Задра Г., Батиста Дж. Л., Лода М. Анализ двойной роли AMPK в раке: от экспериментальных до исследований на людях. Мол Рак Рез. 2015;13:1059–1072. [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Иган Д.Ф., Шакелфорд Д.Б., Михайлова М.М., Гелино С., Конц Р.А., Майр В. и соавт. Фосфорилирование ULK1 (hATG1) с помощью AMP-активируемой протеинкиназы связывает восприятие энергии с митофагией. Наука. 2011; 331:456–461. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Наука. 2011; 331:456–461. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Чон С.М., Чандел Н.С., Хэй Н. AMPK регулирует гомеостаз NADPH, способствуя выживанию опухолевых клеток во время энергетического стресса. Природа. 2012; 485: 661–665. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Choudhury Y., Yang Z., Ahmad I., Nixon C., Salt I.P., Leung H.Y. AMP-активированная протеинкиназа (AMPK) как потенциальная терапевтическая мишень, независимая от передачи сигналов PI3K/Akt, при раке предстательной железы. Онсознание. 2014; 1: 446–456. [Бесплатная статья PMC] [PubMed] [Google Scholar]
19. Muaddi H., Chowdhury S., Vellanki R., Zamiara P., Koritzinsky M. Вклад AMPK и p53-зависимой передачи сигналов в ответ на облучение в присутствии метформина. Радиотер Онкол. 2013; 108: 446–450. [PubMed] [Google Scholar]
20. Zhou G., Wang J., Zhao M., Xie T.X., Tanaka N., Sano D., et al. Мутантный p53 с приобретением функции способствует росту клеток и метаболизму раковых клеток посредством ингибирования активации AMPK. Мол Ячейка. 2014;54:960–974. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мол Ячейка. 2014;54:960–974. [Бесплатная статья PMC] [PubMed] [Google Scholar]
21. Ли Д.Х., Ли Т.Х., Юнг Ч.Х., Ким Ю.Х. Вогонин индуцирует апоптоз, активируя сигнальные пути AMPK и p53 в клетках глиобластомы человека. Сотовый сигнал. 2012;24:2216–2225. [PubMed] [Google Scholar]
22. Li QR, Yan XM, Guo L., Li J., Zang Y. AMPK регулирует длину центрального веретена анафазы путем фосфорилирования KIF4A. J Mol Cell Biol. 2018;10:2–17. [PubMed] [Google Scholar]
23. Visnjic D., Dembitz V., Lalic H. Роль модуляторов AMPK/mTOR в терапии острого миелоидного лейкоза. Курр Мед Хим. 2019;26:2208–2229. [PubMed] [Google Scholar]
24. You MK, Kim HJ, Kook JH, Kim HA. Зверобой регулирует пролиферацию и апоптоз в клетках рака молочной железы человека MCF-7, ингибируя AMPK/mTOR и активируя митохондриальный путь. Int J Mol Sci 2018; 19:966. [Бесплатная статья PMC] [PubMed]
25. Bungard D., Fuerth B.J., Zeng P.Y., Faubert B., Maas N.L., Viollet B., et al. Сигнальная киназа AMPK активирует стрессорную транскрипцию посредством фосфорилирования гистона h3B. Наука. 2010;329: 1201–1205. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Наука. 2010;329: 1201–1205. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Zhang C.S., Lin S.C. AMPK способствует аутофагии, способствуя делению митохондрий. Клеточный метаб. 2016;23:399–401. [PubMed] [Google Scholar]
27. Toyama E.Q., Herzig S., Courchet J., Lewis T.L., Jr., Loson O.C., Hellberg K., et al. Активируемая АМФ протеинкиназа опосредует деление митохондрий в ответ на энергетический стресс. Наука. 2016; 351: 275–281. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Jiang Z., Chen X., Chen K., Sun L., Gao L., Zhou C., et al. Ингибирование YAP ресвератролом посредством активации AMPK повышает чувствительность раковых клеток поджелудочной железы к гемцитабину. Питательные вещества. 2016;8:546. [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Fu X., Wan S., Lyu Y.L., Liu L.F., Qi H. Этопозид индуцирует ATM-зависимый митохондриальный биогенез посредством активации AMPK. ПЛОС Один. 2008;3:e2009. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. Luo L., Huang W., Tao R., Hu N., Xiao Z.X., Luo Z. Зависимая от ATM и LKB1 активация AMPK повышает чувствительность раковых клеток к этопозиду. -индуцированный апоптоз. Рак Летт. 2013; 328:114–119. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Luo L., Huang W., Tao R., Hu N., Xiao Z.X., Luo Z. Зависимая от ATM и LKB1 активация AMPK повышает чувствительность раковых клеток к этопозиду. -индуцированный апоптоз. Рак Летт. 2013; 328:114–119. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Toulany M., Mihatsch J., Holler M., Chaachouay H., Rodemann H.P. Опосредованная цисплатином радиосенсибилизация клеток немелкоклеточного рака легкого стимулируется ингибированием АТМ. Радиотер Онкол. 2014; 111: 228–236. [PubMed] [Академия Google]
32. Негрини С., Горгулис В.Г., Халазонетис Т.Д. Геномная нестабильность – развивающийся признак рака. Nat Rev Mol Cell Biol. 2010;11:220–228. [PubMed] [Google Scholar]
33. Zannella V.E., Cojocari D., Hilgendorf S., Vellanki R.N., Chung S., Wouters B.G., et al. AMPK регулирует метаболизм и выживаемость в ответ на ионизирующее излучение. Радиотер Онкол. 2011;99:293–299. [PubMed] [Google Scholar]
34. Sanli T., Storozhuk Y., Linher-Melville K., Bristow R.G., Laderout K. , Viollet B., et al. Ионизирующее излучение регулирует экспрессию AMP-активируемой протеинкиназы (AMPK) в эпителиальных раковых клетках: модуляция клеточных сигналов, регулирующих клеточный цикл и выживание. Радиотер Онкол. 2012;102:459–465. [PubMed] [Google Scholar]
, Viollet B., et al. Ионизирующее излучение регулирует экспрессию AMP-активируемой протеинкиназы (AMPK) в эпителиальных раковых клетках: модуляция клеточных сигналов, регулирующих клеточный цикл и выживание. Радиотер Онкол. 2012;102:459–465. [PubMed] [Google Scholar]
35. Ui A., Ogiwara H., Nakajima S., Kanno S., Watanabe R., Harata M., et al. Возможное участие передачи сигналов LKB1-AMPK в негомологичном соединении концов. Онкоген. 2014; 33:1640–1648. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Xin J., Yang H., Fan N., Zhao B., Ouyang Z., Liu Z., et al. Высокоэффективное поколение биаллельных инбредных мини-свиней с нокаутом GGTA1 с TALEN. ПЛОС Один. 2013;8:e84250. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Исибаши С., Клифф Р., Амайя Э. Высокоэффективные уровни биаллельных мутаций с использованием TALEN в Xenopus tropicalis . Биол открытый. 2012; 1:1273–1276. [Бесплатная статья PMC] [PubMed] [Google Scholar]
38. Liu Y., Luo D. , Zhao H., Zhu Z., Hu W., Cheng C.H. Наследуемые и точные большие геномные делеции некодирующих генов РНК у рыбок данио с использованием TALEN. ПЛОС Один. 2013;8:e76387. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Zhao H., Zhu Z., Hu W., Cheng C.H. Наследуемые и точные большие геномные делеции некодирующих генов РНК у рыбок данио с использованием TALEN. ПЛОС Один. 2013;8:e76387. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Panda S.K., Wefers B., Ortiz O., Floss T., Schmid B., Haass C., et al. Высокоэффективный направленный мутагенез у мышей с использованием TALEN. Генетика. 2013;195: 703–713. [PMC free article] [PubMed] [Google Scholar]
40. Katsuyama T., Akmammedov A., Seimiya M., Hess S.C., Sievers C., Paro R. Эффективная стратегия TALEN-опосредованной инженерии генома Drosophila . Нуклеиновые Кислоты Res. 2013;41:e163. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Wefers B., Panda S.K., Ortiz O., Brandl C., Hensler S., Hansen J., et al. Создание целевых мутантов мыши путем микроинъекции эмбриона мРНК TALEN. Нат Проток. 2013; 8: 2355–2379. [PubMed] [Google Scholar]
42. Ferguson C., McKay M., Harris R.A., Homanics G.E. Крысы с нокаутом Toll-подобного рецептора 4 (Tlr4), полученные инактивацией гена, опосредованной активатором транскрипции эффекторной нуклеазой (TALEN). Алкоголь. 2013; 47: 595–599. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Алкоголь. 2013; 47: 595–599. [Бесплатная статья PMC] [PubMed] [Google Scholar]
43. Ding Q., Lee Y.K., Schaefer E.A., Peters D.T., Veres A., Kim K., et al. Система редактирования генома TALEN для создания моделей заболеваний на основе стволовых клеток человека. Клеточная стволовая клетка. 2013;12:238–251. [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Fernandez-Capetillo O., Chen H.T., Celeste A., Ward I., Romanienko P.J., Morales J.C., et al. Индуцированная повреждением ДНК активация контрольной точки G2-M гистонами h3AX и 53BP1. Nat Cell Biol. 2002; 4: 993–997. [PubMed] [Google Scholar]
45. Декбар Д., Джегго П.А., Лобрич М. Понимание ограничений радиационно-индуцированных контрольных точек клеточного цикла. Crit Rev Biochem Mol Biol. 2011; 46: 271–283. [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. Рупрехт Б., Роесли С., Лемер С., Кастер Б. MALDI-TOF и nESI Orbitrap MS/MS идентифицируют ортогональные части фосфопротеома. Протеомика. 2016;16:1447–1456. [PubMed] [Академия Google]
[PubMed] [Академия Google]
47. Викрамасингхе В.О., Венкитараман А.Р. Процессинг РНК и стабильность генома: причина и следствие. Мол Ячейка. 2016; 61: 496–505. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Харди Д.Г., Шаффер Б.Е., Брюнет А. AMPK: путь восприятия энергии с несколькими входами и выходами. Тенденции клеточной биологии. 2016;26:190–201. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Bergamaschi D., Samuels Y., Jin B., Duraisingham S., Crook T., Lu X. ASPP1 и ASPP2: общие активаторы членов семейства p53. Мол Селл Биол. 2004; 24:1341–1350. [Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Slee E.A., Lu X. Семья ASPP: выбор между жизнью и смертью после повреждения ДНК. Токсикол Летт. 2003; 139:81–87. [PubMed] [Google Scholar]
51. Huang X., Yan J., Zhang M., Wang Y., Chen Y., Fu X., et al. Ориентация на эпигенетические перекрестные помехи как терапевтическая стратегия для EZh3-аберрантных солидных опухолей. Клетка.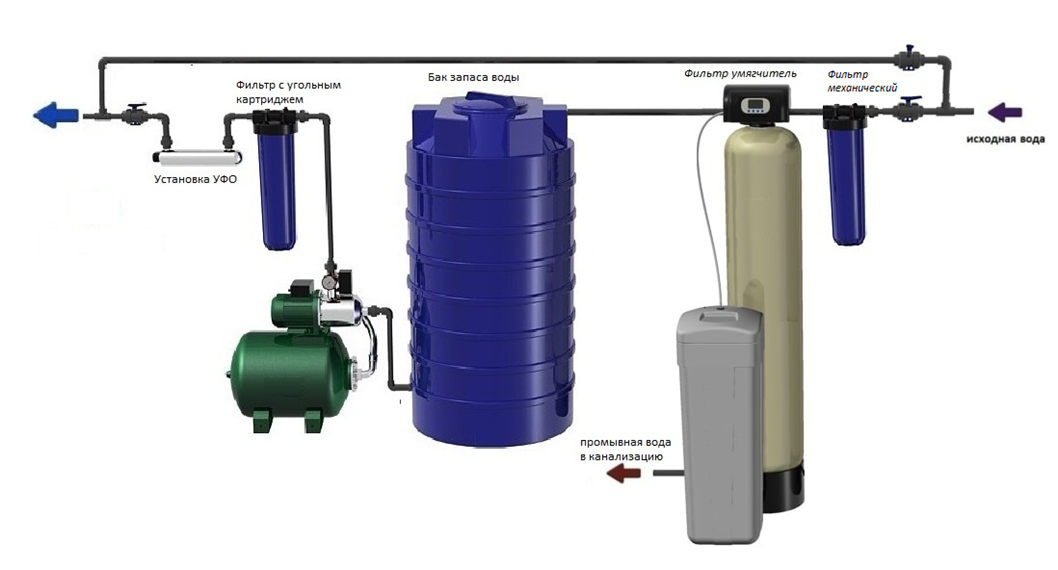 2018; 175:186–199. е19. [PubMed] [Google Scholar]
2018; 175:186–199. е19. [PubMed] [Google Scholar]
52. Накаока С., Сасаки К., Ито А., Накао Ю., Йошида М. Генетически кодируемый зонд FRET для обнаружения внутринуклеосомного гистона h4K9или ацетилирование h4K14 с использованием BRD4, члена семейства BET. ACS Chem Biol. 2016; 11: 729–733. [PubMed] [Google Scholar]
53. Liu Y., Wang X., Zhang J., Huang H., Ding B., Wu J., et al. Структурная основа и связывающие свойства второго бромодомена Brd4 с ацетилированными гистоновыми хвостами. Биохимия. 2008;47:6403–6417. [PubMed] [Google Scholar]
54. Филиппакопулос П., Пико С., Мангос М., Китс Т., Ламберт Дж. П., Барсайт-Лавджой Д. и др. Распознавание гистонов и крупномасштабный структурный анализ семейства бромодоменов человека. Клетка. 2012;149: 214–231. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Choi S., Bakkenist CJ Brd4 защищает хроматин от сигнальных штормов киназы ATM. Научный сигнал. 2013;6:pe30. [Бесплатная статья PMC] [PubMed] [Google Scholar]
56. Zhang C., Su Z.Y., Wang L., Shu L., Yang Y., Guo Y., et al. Эпигенетическая блокада неопластической трансформации бромодоменом и ингибитором белка экстратерминального (ВЕТ) домена JQ-1. Биохим Фармакол. 2016;117:35–45. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Zhang C., Su Z.Y., Wang L., Shu L., Yang Y., Guo Y., et al. Эпигенетическая блокада неопластической трансформации бромодоменом и ингибитором белка экстратерминального (ВЕТ) домена JQ-1. Биохим Фармакол. 2016;117:35–45. [Бесплатная статья PMC] [PubMed] [Google Scholar]
57. Стюарт Х.Дж., Хорн Г.А., Бастоу С., Чевассут Т.Дж. BRD4 связывается с p53 в клетках лейкемии с мутацией DNMT3A и участвует в апоптозе под действием ингибитора бромодомена JQ1. Рак Мед. 2013;2:826–835. [Статья PMC free] [PubMed] [Google Scholar]
58. Dey A., Chitsaz F., Abbasi A., Misteli T., Ozato K. Белок двойного бромодомена Brd4 связывается с ацетилированным хроматином во время интерфазы и митоза. Proc Natl Acad Sci U S A. 2003;100:8758–8763. [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Катайисто П., Валлениус Т., Вахтомери К., Экман Н., Удд Л., Тиайнен М. и соавт. Киназа-супрессор опухолей LKB1 при заболеваниях человека. Биохим Биофиз Акта. 2007; 1775: 63–75. [PubMed] [Google Scholar]
60. Kim J., Kundu M., Viollet B., Guan K.L. AMPK и mTOR регулируют аутофагию посредством прямого фосфорилирования Ulk1. Nat Cell Biol. 2011;13:132–141. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Kim J., Kundu M., Viollet B., Guan K.L. AMPK и mTOR регулируют аутофагию посредством прямого фосфорилирования Ulk1. Nat Cell Biol. 2011;13:132–141. [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. van Veelen W., Korsse S.E., van de Laar L., Peppelenbosch M.P. Долгий и извилистый путь к рациональному лечению рака, связанного с передачей сигналов LKB1/AMPK/TSC/mTORC1. Онкоген. 2011;30:2289–2303. [PubMed] [Google Scholar]
62. Howell J.J., Hellberg K., Turner M., Talbott G., Kolar M.J., Ross D.S., et al. Метформин ингибирует передачу сигналов mTORC1 в печени посредством дозозависимых механизмов, включающих AMPK и комплекс TSC. Клеточный метаб. 2017; 25: 463–471. [Бесплатная статья PMC] [PubMed] [Google Scholar]
63. Иноки К., Ким Дж., Гуан К.Л. AMPK и mTOR в гомеостазе клеточной энергии и лекарственные мишени. Annu Rev Pharmacol Toxicol. 2012;52:381–400. [PubMed] [Google Scholar]
64. Уайтсайд Т.Л. Микроокружение опухоли и его роль в стимулировании роста опухоли. Онкоген. 2008;27:5904–5912. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Онкоген. 2008;27:5904–5912. [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Хайдеггер И., Пирчер А., Пихлер Р. Ориентация на микроокружение опухоли в биологии и терапии почечно-клеточного рака. Фронт Онкол. 2019;9:490. [Бесплатная статья PMC] [PubMed] [Google Scholar]
66. Ханахан Д., Вайнберг Р.А. Отличительные признаки рака: следующее поколение. Клетка. 2011; 144:646–674. [PubMed] [Google Scholar]
67. Reynolds T.Y., Rockwell S., Glazer P.M. Генетическая нестабильность, индуцированная микроокружением опухоли. Рак Рез. 1996;56:5754–5757. [PubMed] [Google Scholar]
68. Xu H., Zhou Y., Coughlan K.A., Ding Y., Wang S., Wu Y., et al. Дефицит AMPKalpha1 способствует клеточной пролиферации и повреждению ДНК за счет снижения p21 в эмбриональных фибробластах мыши. Биохим Биофиз Акта. 2015; 1853: 65–73. [Бесплатная статья PMC] [PubMed] [Google Scholar]
69. Rashid A, Liu C, Sanli T, Tsiani E, Singh G, Bristow RG, et al. Ресвератрол усиливает реакцию клеток рака предстательной железы на ионизирующее излучение. Модуляция путей AMPK, Akt и mTOR. Радиат Онкол 2011;6:144. [Бесплатная статья PMC] [PubMed]
Модуляция путей AMPK, Akt и mTOR. Радиат Онкол 2011;6:144. [Бесплатная статья PMC] [PubMed]
70. Samuels-Lev Y., O’Connor D.J., Bergamaschi D., Trigiante G., Hsieh J.K., Zhong S., et al. Белки ASPP специфически стимулируют апоптотическую функцию p53. Мол Ячейка. 2001; 8: 781–794. [PubMed] [Google Scholar]
71. Вивес В., Су Дж., Чжун С., Ратнаяка И., Сли Э., Голдин Р. и соавт. ASPP2 является гаплонедостаточным супрессором опухоли, который взаимодействует с p53 для подавления роста опухоли. Гены Дев. 2006; 20:1262–1267. [Бесплатная статья PMC] [PubMed] [Google Scholar]
72. Мори Т., Окамото Х., Такахаши Н., Уэда Р., Окамото Т. Аберрантная сверхэкспрессия мРНК 53BP2 в клеточных линиях рака легких. ФЭБС лат. 2000;465:124–128. [PubMed] [Академия Google]
73. Ao Y., Rohde L.H., Naumovski L. p53-взаимодействующий белок 53BP2 ингибирует клоногенное выживание и повышает чувствительность клеток к доксорубицину, но не к паклитаксел-индуцированному апоптозу. Онкоген.